
Федеральное Государственное Учреждение Здравоохранения цмсч №165 Федерального Медико-Биологического Агентства справочное методическое пособие
| Вид материала | Методическое пособие |
| Гормоны и гормоноиды Продуцируемые информоны Схема . Синтез катехоламинов. Схема . Метаболизм и мочевая экскреция катехоламинов. МАО – моноаминоксидаза; КОМТ – катехол-О-метилтрансфераза. Значение тонуса |
- Работа выполнена в фгу «Государственный научный центр лазерной медицины» Федерального, 275.62kb.
- Утверждено, 220.09kb.
- «Научно-исследовательский институт детских инфекций Федерального медико-биологического, 263.32kb.
- Комплексный подход к диагностике и хирургическому лечению заболеваний щитовидной железы, 1223.26kb.
- «Оценка эффективности и безопасности структурно-резонансной терапии у неврологических, 1777.93kb.
- «Санкт-Петербургский институт усовершенствования врачей-экспертов Федерального медико-биологического, 361.07kb.
- Федеральное медико-биологическое агентство, 6378.43kb.
- На правах рукописи, 423.43kb.
- Государственное образовательное учреждение высшего профессионального образования московский, 1119.17kb.
- Государственное образовательное учреждение высшего профессионального образования московский, 1335.69kb.

Гормоны и гормоноиды
| Синтезирующий орган | Продуцируемые информоны | Мишень информационного влияния | Направление воздействия | | ||||
| Эпифиз | Альдостерон-стимулирующий гормон (АСГ) | Гломерулярный слой коры надпочечников | Стимуляция (+) | | ||||
| Мелатонин | Гипоталамические либерины (особенно СТГ, ЛГ) | Ингибирование (–) | | |||||
| Аденогипофиз (передняя и средняя доли гипофиза) | Пропио- мелано- кортин | -липотропин | АКТГ | + | | |||
| -липотропин | Пищевое поведение и жировой обмен | регуляция | | |||||
| альфа-меланоцитстимулирующий гормон (-МСГ) | Меланин специальных клеток на периферии | + | | |||||
| -МСГ | Альдостерон гломерулярного слоя надпочечников | + | | |||||
| эндорфины | Околоводопроводное (сильвиев водопро- вод), серое вещество и большое ядро шва — опиоидные рецепторы | + | | |||||
| адренокортикотропный гормон (АКТГ) | Кора надпочечников (ГКС, альдостерон, андростендион) | + | | |||||
| Меланоциты | + | | ||||||
| Соматотропный гормон (СТГ) | Инсулиноподобный фактор роста (ИФР) тканей | + | | |||||
| Глюконеогенез в печени | + | | ||||||
| Липолиз в адипоцитах | | | ||||||
| Пролактин (лютеотропный гормон – ЛТГ) | Молочные железы | + | | |||||
| Желтое тело яичника | + персистенция | | ||||||
| Имплантация оплодотворенной яйцеклетки | + | | ||||||
| Гонадолиберин гипоталамуса | – | | ||||||
| Тиреотропный гормон (ТТГ) | Щитовидная железа | + | | |||||
| Лютеинизирующий гормон (ЛГ) лютропин гонадотропин ГСИК (гормон стимуляции интерстициальных клеток яичка) | Овуляция и образование желтого тела | + | | |||||
| Интерстициальные клетки Лейдига в яичках | + тестостерон | | ||||||
| “Тека” коркового слоя яичников | + андрогены | | ||||||
| Сетчатая зона коры надпочечников | + андростендион | | ||||||
| Фолликулостимулирующий гормон (ФСГ) фоллитропин | Рост и созревание фолликулов в яичниках | + | | |||||
| Клетки Сертоли яичка | + развитие сперматозоидов | | ||||||
| Гранулезные клетки яичника (переход андрогенов в эстрогены) | + | | ||||||
| Нейрогипофиз (задняя доля гипофиза) | Антидиуретический гормон (АДГ) вазопрессин | Гиалуронидаза нефрона | + | | ||||
| Сосуды (в т.ч. V.portae) | + | | ||||||
| Окситоцин | Желтое тело яичника | + | | |||||
| Матка | + | | ||||||
| Молочные железы | + | | ||||||
| Семенные канальцы | + | | ||||||
| Гипоталамус | Адреногломерулотрофин | АСГ эпифиза | + | | ||||
| Кортиколиберин | АКТГ аденогипофиза | + | | |||||
| Нейрофизин I | Предшественник окситоцина нейрогипофиза | | | |||||
| Нейрофизин II (проАДГ) | Предшественник АДГ нейрогипофиза | | | |||||
| Фоллиберин | ФСГ аденогипофиза | + | | |||||
| Гонадолиберин (лютеин-рилизинг-фактор) | ЛГ аденогипофиза | + | | |||||
| Тиролиберин | ТТГ и ЛТГ аденогипофиза | + | | |||||
| Соматотропин рилизинг-фактор | СТГ аденогипофиза | + | | |||||
| Соматостатин рилизинг-фактор | СТГ, ТТГ, ЛТГ аденогипофиза | – | | |||||
| Меланоцитстимулирующий гормон | -МСГ аденогипофиза | + | | |||||
| Меланостатин | -МСГ аденогипофиза | – | | |||||
| Пролакто- статин | дофамин | ЛТГ аденогипофиза | – | |||||
| лейэнкефалин | ЛТГ аденогипофиза | – | ||||||
| метэнкефалин | ЛТГ аденогипофиза | – | ||||||
| Щитовидная железа | Тироксин | Все виды метаболизма | + | | ||||
| Кальцитонин | Снижение уровня Ca2+ в гомеостазе с участием разных органов | | | |||||
| Паращитовидные железы | Паратирин (ПТГ) | Повышение уровня Ca2+ в гомеостазе с участием разных органов | | | ||||
| Вилочковая железа | Тимозины; Тимопоэтины | Т-лимфоциты | + | | ||||
| Надпочечники мозговой слой | Адреналин | Адренергические рецепторы | + | | ||||
| Надпочечники корковый слой | Альдостерон | Na+/K+ АТФаза натриевых каналов нефрона | + | | ||||
| Глюкокортикостероиды | Все виды метаболизма | различно | | |||||
| Андростеновые стероиды | Предшественники тестостерона и эстрадиола | | | |||||
| Прегнановые стероиды | Предшественники половых гормонов и альдосте рона | | | |||||
| Почки | Эритропоэтин | Эритропоэз | + | | ||||
| Поджелудочная железа | Инсулин | Все виды метаболизма | | | ||||
| Глюкагон | Все виды метаболизма | | | |||||
| Печень | Ангиотензиноген | Предшественники ангиотензина I | | | ||||
| Соматомедин | Антагонист СТГ | | | |||||
| Вариант витамина Д3 (холестановый стероид) | Обмен Ca2+ | | | |||||
| Половые железы мужские | Андростендион | Предшественники тестостерона и эстрадиола | | | ||||
| Тестостерон | Сперматогенез | + | | |||||
| Ингибин | Вилочковая железа | – | | |||||
| Активин | ФСГ аденогипофиза | | | |||||
| Половые железы женские | Эстрогены | Эндометрий и многие органы у женщин (в т.ч. у мужчин — на структурно-функциональное состояние многих органов) | | | ||||
| Прогестерон | Подготовка и поддержание беременности | | | |||||
| Фолликулостатин (ингибин) | ФСГ гипофиза | | | |||||
| Плацента | Эстрогены | Многие ткани | | | ||||
| Прогестины | Многие ткани | | | |||||
| Хориогонический гонадотропин | Эффект, близкий ТТГ | | | |||||
| Релаксин | Связочный аппарат симфиза | – | | |||||
| Пролактин (плацентарный лактоген) | Желтое тело яичника Молочные железы | | | |||||
По многим функциональным признакам вещество Р (аналогично нейрокинам А и В) следует отнести к группе тахикининов: сходный спектр физиологических функций, общая система рецепторов, родственные признаки структуры предшественников. Обладает широким спектром физиологической активности: изменение артериального давления крови, капиллярной проницаемости, сокращение гладкой мускулатуры, секретогенное действие, высвобождение пролактина и пищеварительных гормонов. В последнее время интенсивно изучается роль вещества Р и его аналогов в регуляции центральных процессов – порога болевого воздействия, обучения, сна, устойчивости к стрессу. В головном мозге вещество Р участвует в процессах, связанных с функцией другого нейрорегулятора – допамина. При повреждении допаминэргических волокон выявлено снижение экспресии мРНК, кодирующих образование вещества Р, энкефалинов, динорфина.
В синовиальной жидкости при артрите отмечается высокий уровень вещества Р. Повышенный уровень вещества Р в синовиальной жидкости вызывает усиление продукции синовиоцитами коллагеназы и свободных радикалов кислорода. Такое местное выделение может быть достаточно интенсивным и приводить к локальному уменьшению вещества Р в нервных терминалях. Кроме того, микроокружение в синовиальной ткани содержит много молекул, способных стимулировать хемочувствительные нервы. К таким непосредственно стимулирующим агентам относят брадикинин, гистамин, серотонин. Их действие усиливается веществом Р и воспалением. Эти прямые химические стимуляторы, возможно, ответственны за возникновение боли в покое. Воспалительные медиаторы (ИЛ-1, ФНО-α) приводят к выделению нейропептидов из сенсорных окончаний (увеличивается уровень субстанции Р) и ускоряется деградация хряща. Вещество Р стимулирует тучные клетки, которые в результате могут выделять вазоактивный интестинальный пептид и фактор роста нервов.
Нейротензин представляет собой тетрадекапептид; последовательность 5 из 9 аминокислот в вазопрессине и 5 из 10 аминокислот в ГнРГ идентична или сходна с таковой в нейротензине.
В гипоталамусе присутствует 30% от его общего содержания в мозге; остальное количество находится в среднем мозге и стволе, но самая высокая концентрация характерна именно для гипоталамуса и гипофиза (по данным иммунологических определений). Общее содержание нейротензина в мозге в 10 раз меньше,, чем в экстрактах тонкого кишечника. Мозговая и кишечная форма обладают, по-видимому, химическим сходством.
Кроме влияния на кровообращение (расширение сосудов и гипотензия), нейротензин вызывает и резкую гипергликемию у гипофиз- и адреналэктомированных или получавших морфин, феноксибензамин или пропранолол (анаприлин) животных. Его гипергликемический эффект обусловлен в основном стимуляцией секреции глюкагона и угнетением секреции инсулина, что опосредуется отчасти гормонами мозгового слоя надпочечников, а отчасти — гистамином.
Нейротензин принимает участие и в стимуляции секреции СТГ, пролактина, ТТГ и АКТГ и может аналогичным образом влиять на секрецию гонадотропинов. Поскольку эти эффекты наблюдаются после внутривенной инъекции нейротензина, не исключено, что они обусловлены сосудорасширяющим действием.
В отличие от этого, введение нейротензина в желудочки головного мозга угнетает секрецию СТГ, пролактина и ТТГ, а также вызывает гипотермию.
Церебролизин (цереброл) представляет собой концентрат, содержащий низкомолекулярные биологически активные нейропептиды (лейэнкефалин, метэнкефалин, нейротензин, вещество Р, β-эндорфин и др.) с молекулярным весом, не превышающим 10 000 дальтон (15 %), и свободные аминокислоты (85 %).
Церебролизин является опосредованным ингибитором Са2+-зависимой протеазы кальпаина и обеспечивает активацию синтеза эндогенных кальпостатинов. Эффект Церебролизина на систему кальпаин — кальпостатин является многогранным и опосредованным через систему внутриклеточных антиоксидантов. Он зависит от присутствия в препарате нейропептидов и металлолигандных комплексов, которые выступают конкурентными антагонистами обратимой Cа2+-зависимой активации кальпаина и стабилизаторами цитоскелета нейронов (Wronski с соавт., 2000). Церебролизин обладает способностью нормализовывать пластический обмен в пресинаптических окончаниях и предотвращать нарушения продукции белка — предшественника амилоида. Церебролизин тормозит активацию микроглии in vivo и in vitro , что способствует торможению иммуновоспалительных нарушений в мозге на последних этапах нейродегенеративной ремодуляции через торможение высвобождения цитокинов ИЛ-1, ИЛ-6. Данные современной нейрохимии свидетельствуют о наличии у Церебролизина свойств мембранопротектора, способного регулировать кальциевый гомеостаз, уменьшать нейротоксическое действие повышенных концентраций возбуждающих аминокислот (глутамата).
Церебролизин улучшает транспорт глюкозы через ГЭБ (продукция GLUT1) , тем самым увеличивая число жизнеспособных нейронов и продлевая время выживаемости последних после ишемии и гипоксии; способен ингибировать образование. ОН-радикалов при экспериментальной ишемии (у мышей). Была доказана способность Церебролизина защищать митохондрии нейронов от повреждающего воздействия лактат-ацидоза.).
Церебролизин тормозит апоптоз нейронов и улучшает рост дендритов и аксонов .
Церебролизин содержит макроэлементы (МаЭ) и эссенциальные микроэлементы (МЭ), проявляет витаминную активность тиамина (витамина В1), фолиевой кислоты , цинкобаламина, витамина Е, содержит до 100 короткоцепочечных пептидов , в том числе глутатион и тиролибериновые мотивы. В эксперименте Церебролизин повышает в гипоталамусе, центральной коре, обонятельных луковицах уровень Li, B, Se.
Церебролизин является непрямым блокатором кальпаина и действует через систему внутриклеточных антиоксидантов, что зависит от присутствия в составе препарата нейропептидов и металлолигандных комплексов, которые выступают конкурентными антагонистами Cа2+-зависимой активации кальпаина и деградации цитоскелета нейронов при нейродегенеративных и ишемических заболеваниях мозга.
Церебрамин получен из коры головного мозга крупного рогатого скота и представляет собой комплекс белков и нуклеопротеидов, обладающих избирательным действием на клетки головного мозга, способствующих ускорению восстановления функций мозга и улучшению памяти.
Дельтаран представляет собой композицию на основе синтетического нонапептида — аналога одного из важнейших природных эндогенных пептидов--- регуляторного пептида дельта-сна (ДСИП: дельта-сон индуцирующий пептид) со стабилизирующим клеточный гомеостаз действием (капли в нос).
Основным механизмом действия препарата является возмещение возникающего при психоэмоциональном или системном стрессе, токсических воздействиях, старении организма дефицита ДСИП, который защищает нейроны от разрушения. Модулирующее, улучшающее скоординированность всех звеньев ЦНС влияние ДСИП реализуется через классические нейромедиаторные системы: адренергические, серотонин- и ГАМКергические. Защищая центральную нервную систему от повреждающих воздействий, Дельтаран способен радикально менять течение большого количества заболеваний, так как стрессовые реакции, сопровождающие любой патологический процесс или экстремальное внешнее воздействие, приводят к разрушению нейронов, нарушают работу ЦНС и, следовательно, способность головного мозга поддерживать гомеостаз. Может применяться при алкогольной абстиненции.
Семакс - синтетический аналог фрагмента адренокортикотропного гормона из семи аминокислот (метионил-глутамил-гистидил-фенилаланил-пролил-глицил-пролин), синтетический нейропептид, обладающий ноотропными и нейропротекторными свой-ствами, полностью лишенный гормональной активности.Семакс ограничивает активность NMDA-рецепторов и тем самым снижал развитие глутаматной эксайтотоксичности. Семакс устраняет дисбаланс про- и противовоспалительных цитокинов и повышает содержание нейротрофических факторов (NGF-β1, BDNF). Препарат влияет на процессы, связанные с формированием памяти и обучением. Семакс усиливает избирательное внимание в момент восприятия и анализа информации, улучшает закрепление памятного следа и его хранение, что находит применение как у здоровых лиц, так и у пациентов с различной неврологической патологией. Препарат улучшает адаптацию организма к гипоксии, церебральной ишемии, наркозу и другим повреждающим воздействиям.
Селанк - синтетический аналог эндогенного пептида тафтцина; треонил-лизил-пролил-аргинил-пролил-глицил-пролин-диацетат.
Обладает оригинальным механизмом нейроспецифического действия на центральную нервную систему, связывается со специфическими рецепторами на мембранах нервных клеток.
Оказывает влияние на обмен моноаминов в эмоциогенных структурах мозга (гипоталамус, диэнцефалон, кора полушарий) и активность мозговых ферментов тирозин- и триптофангидроксилазы. Проявляет тропность к серотонинергической системе, нормализуя уровень серотонина мозга в условиях экспериментально вызванного его снижения. Селанк стабилизирует процессы возбуждения и торможения в головном мозге и повышает устойчивость нейронов коры полушарий к функциональным нагрузкам высокой интенсивности.
В спектре фармакологического действия препарата преобладает анксиолитический (противотревожный) эффект со стимулирующим (активирующим) компонентом. Препарат не обладает гипноседативными и миорелаксантными свойствами. Положительно влияет на мнестические и когнитивные функции мозга, в том числе при их нарушении. Активирует процессы обучения, памяти, анализа и воспроизведения информации, улучшает параметры внимания и краткосрочной памяти. Повышает мотивационную устойчивость и адекватность адаптивного поведения. Селанк обладает вегетотропным действием, улучшает вегетативное обеспечение деятельности в условиях эмоционального напряжения, оказывает оптимизирующее влияние на адаптационный резерв организма. Селанк не выявляет нежелательного побочного и токсического действия при 200-300-кратном увеличении дозы по сравнению с ED50. He обладает эмбриотоксическим, тератогенным, аллергизирующим, местнораздражающим действием, не обнаруживает мутагенных свойств. У препарата отсутствуют нежелательные отдаленные эффекты. Селанк не вызывает явлений лекарственной зависимости.
Кортексин представляет собой лиофилизат, полученный путём уксуснокислой экстракции из коры головного мозга крупного рогатого скота или свиней, содержащий низкомолекулярные активные нейропептиды, молекулярный вес которых не превышает 10 000 дальтон, достаточный для проникновения через ГЭБ. Обладает тканеспецифическим многофункциональным действием на головной мозг, что проявляется в метаболической регуляции, нейропротекции, функциональной нейромодуляции, нейротрофической активности. Повышает эффективность энергетического метаболизма клеток мозга, улучшает внутриклеточный синтез белка.
Блокирует процессы свободнорадикального окисления. Препарат устраняет дисбаланс тормозных и возбуждающих аминокислот, обладает умеренным GABA-ергическим действием.Препарат оказывает положительное действие при нарушении когнитивных функций, улучшает концентрацию внимания, кратковременную память, способность к обучению, ускоряет восстановление функций головного мозга после стрессорных воздействий, регулирует уровень серотонина и дофамина. Стимулирует процесс умственной деятельности, не оказывая избыточного активирующего влияния, восстанавливает биоэлектрическую активность головного мозга.Стимулирует репаративные процессы в головном мозге. Регулирует соотношение аминокислот и процессы торможения и возбуждения в мозге; он восстанавливает биоэлектрическую активность головного мозга.
Нейропептид Y состоит из 36 аминокислот. Высокие концентрации пептида обнаруживают в мозге и периферической нервной системе. Нейропептид Y обнаруживают, помимо постганглионарных симпатических волокнах, также в мегакариоцитах и тромбоцитах. Действие нейропептида Y связано с угнетением выделения трансмиттера из окончаний симпатических и парасимпатических нервов, а также с постсинаптическим действием (вазоконстрикция). Существует множество подтипов рецепторов к нейропептиду Y, в том числе пресинаптический Y1 и постсинаптический Y2-рецепторы. Центральные эффекты нейропептида Y - гипотензия, гипотермия, угнетение дыхания. Периферические эффекты - сужение мозговых сосудов, гипертензия. Нейропептид постганглионарных симпатических нервов был обнаружен в синовиальной ткани. Повышенные концентрации нейропептида Y в воспалительной синовиальной жидкости свидетельствуют об активном вовлечении симпатического компонента. Нейропептид Y присутствует в некоторых секретомоторных нейронах нервной системы кишечника и может угнетать секрецию воды и электролитов в кишечнике.
CGRP - один из 3-х продуктов гена кальцитонина. Состоит из 37 аминокислот. В больших количествах обнаруживают в С-клетках щитовидной железы. Пептид широко представлен в ЦНС и ПНС, сердечно-сосудистой системе, ЖКТ и мочеполовой системе. При введении в ЦНС вызывает гипотензию, тахикардию, гипертензию. Кокальцигенин является одним из наиболее мощных вазодилататоров.
Пармидин (Пирикарбат).Ангиопротектор, восстанавливает нарушенную микроциркуляцию. Ингибирует брадикинин и калликреин, обладает противовоспалительной активностью, уменьшает проницаемость сосудистой стенки. Препятствует агрегации тромбоцитов, стимулирует фибринолиз, препятствует отложению атерогенных липидов в сосудистой стенке. Уменьшает отек эндотелиальных клеток, способствует обратному развитию атеросклеротических изменений. Антибрадикининовыми свойствами обусловлено прекращение зуда, регрессия эритемы при атопическом дерматите.
Ноопепт ( этиловый эфир N-фенилацетил-L-пролилглицина). Улучшает способность к обучению и память, действуя на все фазы процесса: начальную обработку информации, консолидацию, извлечение. Препятствует развитию амнезии, вызванной электрошоком, блокадой центральных холинергических структур, глутаматергических рецепторных систем, лишением парадоксальной фазы сна. Нейропротекторное действие препарата проявляется в повышении устойчивости мозговой ткани к повреждающим воздействиям (травма, гипоксия, электросудорожное или токсическое действие) и ослаблении степени повреждения нейронов мозга. Препарат уменьшает объем очага на тромботической модели инсульта и предупреждает гибель нейронов в культуре ткани коры головного мозга и мозжечка, подвергнутых действию нейротоксических концентраций глутамата, свободно-радикального кислорода. Оказывает антиоксидантное действие, блокирует потенциал-зависимые кальциевые каналы нейронов, ослабляя нейротоксическое действие избыточного кальция. Улучшает реологические свойства крови, обладая антиагрегантными, фибринолитическими, антикоагулянтными свойствами. Ноотропное действие препарата связано с образованием циклопролилглицина, аналогичного по структуре эндогенному циклическому дипептиду, обладающему антиамнестической активностью, а также с наличием холинопозитивного действия. Увеличивает амплитуду транскаллозального ответа, облегчая ассоциативные связи между полушариями головного мозга на уровне кортикальных структур. Способствует восстановлению памяти и других когнитивных функций, нарушенных в результате повреждающих воздействий - травма мозга, локальная и глобальная ишемия, пренатальные повреждения (алкоголь, гипоксия).Обладает вегетонормализующим действием, способствует уменьшению головных болей, ортостатических нарушений, тахикардии.
Норадреналин
Адренорецепторы подразделяются на альфа и бета-рецепторы . При этом существуют альфа1А , альфа1В- и альфа1С-рецепторы (ранее именовавшиеся соответственно альфа1а, альфа1в и альфа1А), альфа2А -, альфа2В - и альфа2С-, а также бета1-, бета2- и бета3-рецепторы. Все альфа1-рецепторы стимулируют фосфоинозитидный обмен , все альфа2рецепторы ингибируют аденилатциклазу , а все бета-рецепторы ее активируют. Кроме того, для альфа2Арецепторов показано зависимое от G-белков активирование К-каналов, а для альфа2А- и альфа2В-рецепторов G-белок зависимое ингибирование Cа-каналов.
Все адренергические рецепторы имеют классическую для рецепторов, сопряженных с G-белками, структуру - 7 трансмембранных доменов с N-концом полипептида снаружи, а с С-концом - внутри клетки. Размеры этих рецепторов следующие: альфа1А, альфа1В, альфа1С, альфа2А, альфа2В, альфа2С, бета1, бета2 и бета3 аминокислотных остатка. Селективный антагонист альфа1-рецепторов- празозин , альфа2-рецепторов - иохимбин и раувольфцин, а бета-рецепторов - пропранолол .
Мозговой слой надпочечника и симпатическая нервная система являются производными нервного гребешка, т.е. имеют нейроэктодермальное происхождение и служат местом образования катехоламинов, к которым относят дофамин, норадреналин и адреналин. Биосинтез этих низкомолекулярных веществ происходит в хромаффинных клетках мозгового слоя надпочечника, ЦНС и адренергических симпатических волокнах постганглионарных нейронов. Катехоламины являются нейротрансмиттерами, которые опосредуют функцию ЦНС и симпатической нервной системы, принимая основное участие в регуляции сердечно-сосудистой системы. Исходным продуктом для образования катехоламинов является тирозин, который с помощью ряда соединений превращается в адреналин (схема).
Лекарства, стимулирующие вегетативные постганглионарные нейроны. И парасимпатические, и симпатические преганглионарные нейроны в окончаниях своих аксонов секретируют ацетилхолин, который стимулирует постганглионарные нейроны. Более того, введенный в кровь ацетилхолин может также стимулировать постганглионарные нейроны обеих систем, вызывая в организме одновременно и симпатические, и парасимпатические эффекты.
Никотин является другим средством, способным стимулировать постганглионарные нейроны так же, как ацетилхолин, поскольку все мембраны этих нейронов содержат ацетилхолиновые рецепторы никотинового типа. В связи с этим лекарства, стимулирующие постганглионарные нейроны, называют никотиновыми. Некоторые другие лекарства, например метахолин, действуют на оба типа рецепторов — и никотиновые, и мускариновые, тогда как пилокарпин действует только на мускариновые рецепторы.
Никотин возбуждает и симпатические, и парасимпатические постганглионарные нейроны одновременно. В результате возникает сильное сужение сосудов органов брюшной полости и конечностей и одновременно развиваются парасимпатические эффекты, например увеличение активности желудочно-кишечного тракта и иногда — замедление сердцебиений.
Ганглиоблокаторы. Многие важные лекарственные препараты блокируют проведение возбуждения от вегетативных преганглионарных нейронов к постганглионарным нейронам. К ним относят ион тетраэтиламмония, ион гексаметония и пентолиниум. Эти лекарства блокируют стимуляцию постганглионарных нейронов ацетилхолином и в симпатической, и в парасимпатической системах одновременно. Их часто используют для подавления симпатической активности и редко — для блокады парасимпатической активности, поскольку их действие, выключающее симпатическую активность, обычно в значительной мере перекрывает эффекты парасимпатической блокады. Ганглиоблокаторы снижают артериальное давление у больных с гипертензией, но эти лекарства не очень полезны клинически, поскольку их эффекты трудно контролировать.
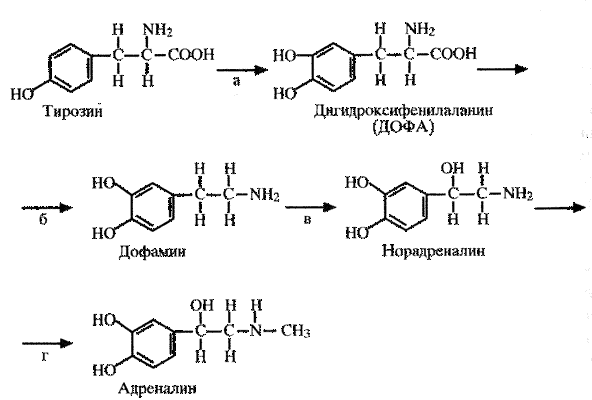
Схема . Синтез катехоламинов.
Вначале происходит гидроксилирование тирозина с образованием дигидроксифенилаланина (ДОФА). Он является предшественником катехоламинов, не обладает биологической активностью, но легко проникает через гематоэнцефалический барьер. Образование ДОФА происходит при участии фермента тирозингидроксилазы (а), которая выявляется в мозговом слое надпочечника, ЦНС и тканях, иннервируемых симпатический нервной системой. Активность тирозингидроксилазы и гидроксилирование тирозина являются основным звеном в биосинтезе катехоламинов, лимитирующим его скорость.
Накопление фенилаланина и его метаболитов угнетает активность тирозингидроксилазы, поэтому при фенилкетонурии синтез катехоламинов снижен. Посредством ДОФА-декарбоксилазы (б) ДОФА превращается в дегидроксифенилэтиламин (дофамин), который при участии дофамин-b-оксидазы (в) и норадреналин-N-метилтрансферазы (г) превращается в норадреналин, а затем в адреналин.
Установлено, что гидроксилирование тирозина с образованием ДОФА происходит в митохондриях хромаффинных клеток. Декарбоксилирование ДОФА и образование дофамина осуществляется в цитозоле клетки, где в растворенном виде присутствуют ДОФА-декарбоксилаза и другие ферменты, необходимые для этого этапа биосинтеза катехоламинов. Дофамин попадает в гранулы клеток или терминали аксонов и в присутствии дофамин-b-оксидазы превращается в норадреналин. Далее норадреналин снова выходит в цитоплазму и, превратившись в адреналин, повторно поглощается гранулами.
Катехоламины в хромаффинных клетках локализуются в гранулах, которые служат резервуаром, местом их биосинтеза и высвобождения. Кроме катехоламинов, гранулы содержат липиды, нуклеотиды (АТФ), белки, ионы Са2+ и Mg2+. В гранулах мозгового слоя надпочечников содержится 80% адреналина и 20% норадреналина. Секреция катехоламинов осуществляется путем экзоцитоза; при этом содержание гранул “изливается” во внеклеточное пространство.
Гранулы выполняют следующие специфические функции: поглощают дофамин из цитозоля клетки и конвертируют его в норадреналин, являются местом “складирования” адреналина и норадреналина, предохраняют их от воздействия моноаминоксидазы и разрушения и в ответ на нервную стимуляцию высвобождают катехоламины в крови. При этом гранулы функционируют как тканевые буферные системы для катехоламинов; эту их функцию можно сравнить с функцией транспортных белков сыворотки крови для тироидных гормонов и кортикостероидов.
В окончаниях симпатических нервных волокон выявляются гранулы, содержащие лишь норадреналин. Аналогичные гранулы обнаружены и в ганглиях симпатической нервной системы. Норадреналин выявлен в головном и спинном мозге, наибольшая концентрация – в области гипоталамуса. Содержание адреналина в этих областях незначительно. Около 80% содержащегося здесь норадреналина локализуется в синаптосомах и нервных окончаниях. Следует отметить, что около 50% катехоламинов, содержащихся в области гипоталамуса и других базальных ганглиях головного мозга, приходится на дофамин.
Высвобождение катехоламинов как из мозгового слоя надпочечников, так и из окончаний симпатической нервной системы происходит под влиянием таких физиологических стимуляторов, как стресс, физическая и психическая нагрузка, повышение уровня инсулина в крови, гипогликемия, гипотония и др. Высвобождение катехоламинов происходит при участии ионов Са2+, который поступает в клетку или в окончания симпатической нервной системы. Поступающие в кровь катехоламины достигают периферических тканей, где накапливаются или метаболизируются прямо пропорционально симпатической иннервации тканей.
Инактивация катехоламинов происходит с участием двух ферментных систем катехол-О-метилтрансферазы (КОМТ) и моноаминоксидазы (МАО). КОМТ является внутриклеточным ферментом, который локализуется в цитоплазме. Считается, что около 50% КОМТ находится в синаптосомах центральной и периферической нервной системы, а остальная часть (50-55%) приходится на другие органы – печень, почку, кишечник, селезенку, слюнные железы, аорту, матку, жировую ткань, эритроциты.
МАО широко представлена в тканях организма (печень, почки, желудок, кишечник, нервная ткань, головной мозг, сердце, половые железы, надпочечники, тромбоциты) и локализуется на наружной мембране митохондрий. В мозговом веществе надпочечников большая часть МАО располагается в митохондриях и лишь незначительное ее количество выявляется в гранулах хромаффинных клеток.
Выявлены два изофермента: МАО-А и МАО-В. МАО-А – фермент нервной клетки, дезаминирующей серотонин, адреналин и норадреналин. МАО-В выявляется в других тканях. На схеме 35 показан обмен катехоламинов, а также их содержание в крови и моче практически здоровых лиц.
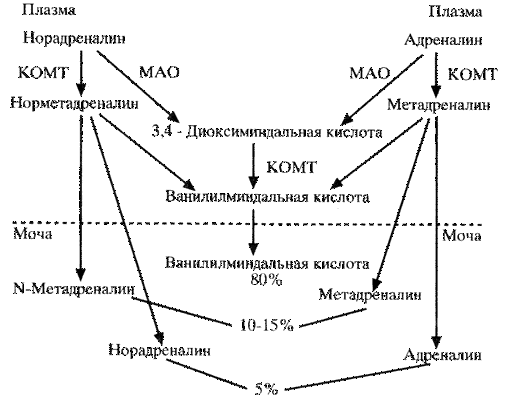
Схема . Метаболизм и мочевая экскреция катехоламинов. МАО – моноаминоксидаза; КОМТ – катехол-О-метилтрансфераза.
Влияние катехоламинов на клеточном уровне опосредуется через адренергические рецепторы, которые можно представить как трансмембранные белки, имеющие 7 трансмембранных фрагментов, а также внеклеточный (аминотерминальный) и внутриклеточный (карбокситерминальный) концы. Эти области адренорецептора ответственны за комплексирование с катехоламинами и хотя имеют очень близкую последовательность аминокислот, но они обладают определенной селективной чувствительностью к различным дофаминовым агонистам, что определяется некоторыми различиями в пятом и седьмом сегменте, которые ответственны за связывание гормона с G-белками. Выше отмечалось, что G-белки состоят из a-, b- и g-субъединиц. Комплексирование гормона с соответствующим рецептором сопровождается диссоциацией a-субъединицы от b- и g-субъединицы. После этого ГТФ замещает ГДФ на a-субъединице и вызывает диссоциацию b- и g-субъединиц. Комплекс a-субъединиц-ГТФ активирует пострецепторные механизмы, приводящие к биологическому эффекту гормона. В результате такого взаимодействия инициируются внутриклеточные процессы, приводящие к физиологическому эффекту. Различают a- и b-адренергические рецепторы, каждый из которых в свою очередь подразделяется на 2 подтипа. a1-адренергические рецепторы опосредуют сосудистые эффекты и сокращения гладких мышц, что проявляется сужением периферических сосудов, расширением зрачка и увеличением потоотделения, тогда как a2-адренергические рецепторы в случае их активирования ингибируют высвобождение норадреналина.
Исследованиями последних лет показано, что a1 в свою очередь подразделяются на a1А, a1В, a1С и a1D, тогда как a2-рецепторы – на a2А, a2В, a2С. Фенилэфрин, метоксамин, циразолин являются агонистом b1-рецептора, тогда как празозин – селективным антагонистом a1-рецептора. Клонидин же относится к агонистам b2-рецептора, а йогимбин и раувольсцин – к селективным антагонистам a2-рецептора. Фентоламин и феноксибензамин действуют подобным образом на оба типа рецепторов. b-Адренергические рецепторы подразделяются на b1, опосредующие прямое стимулирующее влияние на сердце, и b2, которые осуществляют релаксацию гладких мышц матки, бронхов и сосудов, опосредуют гликогенолиз в печени. Агонистом для b-рецепторов является изопротеренол, для b1-рецепторов – добутамин, для b2-рецепторов – прокатерол и тербуталин и для b3-рецепторов BRL37344. Адреналин и норадреналин являются равнозначными агонистами для b1-рецепторов, тогда как норадреналин является слабым агонистом для b2-рецепторов. Антагонистом для b-рецепторов является пропранолол, для b1-рецеторов – бетаксолол, для b2-рецепторов – бутоксамин. b3-Адренергические рецепторы локализуются в жировой ткани и опосредуют липолиз жира.
a-Адренергические рецепторы также подразделяются на 2 вида: a1- и a2-рецепторы. В постсинаптических окончаниях локализуются преимущественно a1-рецепторы, тогда как a2-рецепторы расположены в пресинаптических окончаниях, где они контролируют высвобождение катехоламинов из окончаний симпатических нервов.
Необходимо подчеркнуть, что, несмотря на различное биологическое влияние стимуляции a-, и b-рецепторов на органы и ткани, стимуляция функций желудочно-кишечного тракта и сердца безусловно является результатом активации как a-, так и b-рецепторов.
Катехоламины принимают участие в регуляции секреции гормонов: через b-рецепторный механизм стимулируют высвобождение глюкагона, ренина, гастрина, паратгормона, кальцитонина, инсулина и тироидных гормонов, однако через a-рецепторный механизм угнетают секрецию инсулина.
В последние годы обнаружено наличие третьего типа адренергических рецепторов – дофаминергических, т.е. рецепторов, отвечающих на дофамин (D1, D2, D4, D5 подтипы этих рецепторов), но не на другие катехоламины. Они выявлены в ЦНС (гипоталамус и другие области), в сосудах почек. Гипоталамус модулирует функцию передней доли гипофиза посредством не только гипоталамических гормонов, но и катехоламинов. Было обнаружено, что для стимуляции высвобождения ФСГ и ЛГ и угнетения высвобождения пролактина и СТГ требуются относительно высокие дозы норадреналина и адреналина (5-100 мкг) по сравнению с дофамином (1мкг). Производные лизергиновой кислоты бромокриптин и лерготрил угнетают секрецию пролактина в результате того, что являются агонистами дофаминовых, а правильнее D2-рецепторов,тогда как фенолдопа – агонистом D1-рецепторов. Антагонистом D5 является клозапин. D2- агонисты (бромокриптин и др.) успешно применяются в терапии акромегалии, гиперпролактинемии, болезни Иценко-Кушинга.
На основании проведенных исследований установлено, что a- и b-рецепторы – иммунологически близкие структуры. Под влиянием различных условий возможна транформация a- в b-рецепторы. Это позволило высказать предположение, что a- и b-рецепторы представляют собой аллостерические конформации одной и той же структуры.
b-Адренергическое влияние опосредуется через стимуляцию мембраносвязанного фермента аденилатциклазы, увеличение цАМФ-зависимых протеинкиназ, фосфорилирование специфических белков.
Стимуляция a2-адренергического рецептора сопровождается ингибированием гуанин-нуклеотидного регуляторного белка, что в свою очередь приводит к снижению активности каталитической субъединицы аденилатциклазы и уменьшению образования цАМФ. a1-Адренергическое влияние опосредуется кальций-полифосфоинозидной системой (см. выше). Комплексирование гормона (катехоламинов) с этим рецептором приводит к активизации данной системы: образованию двух мессенджеров – диацилглицерина и инозитол-трифосфата (ИФ3). Последний способствует повышению внутриклеточного кальция, а диацилглицерин активирует специфические протеинкиназы, фосфорилирующие определенные ферменты, что и проявляется различным биологическим эффектом.
Катехоламины влияют на обмен веществ посредством увеличения скорости утилизации энергии и повышения мобилизации энергетических запасов для использования их в тканях. Стимуляция обменных процессов сопровождается повышением образования тепла (термогенез) и при этом увеличивается потребление кислорода. Наличие достаточного количества источников энергии обеспечивается стимуляцией гликогенолиза и липолиза. Дополнительно к прямому действию катехоламины оказывают большое влияние на обмен веществ и через поджелудочную железу: через a- рецепторный механизм снижается секреция инсулина и через b-рецепторный механизм стимулируется высвобождение глюкагона.
При полноценном пищевом рационе в печени человека содержится 100 г (1720 кДж, или 400 ккал) и в скелетных мышцах около 300 г (5160 кДж, или 1200 ккал) гликогена. Влияние катехоламинов на стимуляцию гликогенолиза осуществляется как через активацию аденилатциклазы и цАМФ (b-рецепторы), так и механизмом, связанным с изменением вхождения Са++ в клетку (a-рецепторы). Стимуляция как a, так и b-адренергических рецепторов приводит к увеличению выхода глюкозы из печени. Механизм гликогенолиза, индуцированного катехоламинами в других тканях, менее ясен, но отличается от того, что, описано для печени.
Жировые депо у человека в среднем составляют 15 кг, или 570 500 кДж (135 000 ккал). Катехоламины усиливают липолиз через активацию липазы триглицеридов, что опосредуется через b3-адренергические рецепторы с последующим активированием аденилатциклазы и цАМФ. Имеются сообщения, что стимуляция a-адренергических рецепторов уменьшает скорость липолиза.
Белок в организме представлен в основном в мышечной ткани, где его количество составляет 6 кг (103 200 кДж, или 24 000 ккал). Под влиянием катехоламинов наблюдается протеолиз, хотя увеличивается синтез многих специфических белков. В основном для расхода энергии мобилизуются запасы жиров и углеводов, а белки используются в том случае, если эти источники энергии израсходованы.
Под влиянием катехоламинов стимулируются процессы глюконеогенеза в печени, где для образования глюкозы используются лактат, глицерин и аланин. Эти процессы опосредуются через активацию a-адренергических рецепторов.
Таким образом, биологическое значение катехоламинов в организме велико. Наряду с непосредственным влиянием на обмен веществ катехоламины оказывают опосредованное действие через секрецию других гормонов (СТГ, инсулин, глюкагон, ренин-ангиотензинная система и др.).
Отличительной особенностью автономной нервной системы в сравнении со скелетной (соматической) нервной системой является факт, что для полной активации вегетативных эффекторов требуется лишь небольшая частота стимуляции. В целом достаточно одного нервного импульса в секунду, чтобы поддержать нормальный симпатический или парасимпатический эффект, а полная активация происходит, когда нервные волокна разряжаются с частотой 10-20 имп/сек. Для сравнения: полная активация соматической нервной системы осуществляется при частоте 50-500 имп/сек или более.
В норме симпатическая и парасимпатическая системы постоянно активны, и их базальные уровни активности известны как симпатический тонус и парасимпатический тонус, соответственно.
Значение тонуса состоит в том, что он позволяет одиночной нервной системе как увеличивать, так и снижать активность стимулируемого органа. Например, симпатический тонус в норме держит почти все системные артериолы суженными примерно наполовину от их максимального диаметра. При увеличении степени симпатической стимуляции выше нормы эти сосуды могут сужаться еще больше; наоборот, при снижении стимуляции ниже нормы артериолы могут расширяться. При отсутствии постоянного фонового тонуса симпатическая стимуляции вела бы лишь к сужению сосудов и никогда — к их расширению.