На правах рукописи
| Вид материала | Автореферат |
- Печатная или на правах рукописи, 21.09kb.
- Удк 796/799: 378 , 770.24kb.
- На правах рукописи, 399.58kb.
- На правах рукописи, 726.26kb.
- На правах рукописи, 1025.8kb.
- На правах рукописи, 321.8kb.
- На правах рукописи, 552.92kb.
- На правах рукописи, 514.74kb.
- На правах рукописи, 670.06kb.
- На правах рукописи, 637.26kb.
Результаты исследований и их обсуждение
Полиморфизм нуклеотидных последовательностей генов lcrV и
соответствующих им полипептидов Y. pestis
В настоящей работе используется внутривидовая классификация Y. pestis, приведенная в соответствие с правилами Международного кодекса номенклатуры бактерий. В вид Y. pestis входят два подвида: pestis (включает патогенные для человека биовары intermedium, antiqua, medievalis и orientalis) и microtus (включает непатогенные для человека «полевочьи» биовары altaica, angola, caucasica, hissarica, qinghaiensis, talassica, ulegeica и xilingolensis) (Платонов, 2010).
К моменту начала наших исследований было установлено, что нуклеотидная последовательность гена lcrV штаммов Y. pestis subsp. pestis консервативна, однако секвенирование lcrV гена трех штаммов, принадлежащих к подвиду microtus, выявило вариабельность нуклеотидной последовательности гена данного фактора патогенности (Adair et al., 2000; Song et al., 2004). В связи с этим задачей настоящего раздела исследований явилось изучение полиморфизма нуклеотидных последовательностей гена lcrV и соответствующих им полипептидов у представителей различных подвидов и биоваров Y. pestis, выделенных на территории стран СНГ и Монголии. Особое внимание мы уделили изучению V антигена из штаммов Y. pestis подвида microtus.
Мы провели секвенирование гена lcrV из 73 штаммов разных внутривидовых групп Y. pestis, выделенных в девяти природных очагах стран СНГ, а также на территории Монголии. Причем, 62 из них принадлежало к биоварам altaica, caucasica, hissarica, ulegeica, talassica, которые вместе с биоварами angola, qinghaiensis и xilingolensis входят в состав подвида microtus. Семь последовательностей гена lcrV (штамм subsp. pestis bv. orientalis CO92, штамм subsp. pestis bv. medievalis KIM, штамм subsp. pestis bv. antiqua Antiqua, штамм subsp. pestis bv. orientalis CA88-4125; штамм subsp. microtus bv. caucasica Pestoides F, штамм subsp. microtus bv. angola Angola; штамм subsp. microtus bv. xilingolensis 91001) (Parkhill et al., 2001; Deng et al., 2002; Chain et al., 2006; Eppinger et al., 2009; Eppinger et al., 2010; Song et al., 2004), депонированных в GenBank, анализировали in silico. Таким образом, проанализированы нуклеотидные последовательности гена lcrV и соответствующие им аминокислотные последовательности V антигена из 80 штаммов Y. pestis.
Анализ аминокислотных последовательностей позволил сгруппировать все варианты белка LcrV в пять типов в соответствии с проявлением основного группо-специфичного полиморфизма в четырех «горячих точках» с дополнительным штаммо-специфичным полиморфизмом, выявляемым у представителей отдельных групп. Эти «горячие точки» соответствуют аминокислотным остаткам 18 (K18 → N18), 72 (K72 → R72), 273 (C273 → S273) и 324-326 (S324-G325-K326 → R324) в последовательности LcrV референс-штамма KIM [Deng et al., 2002], а выявленные типы аминокислотных последовательностей были нами описаны следующими формулами:
тип A – N18, R72, S273 и S324-G325- K326;
тип B – N18, R72, C273, и S324-G325-K326;
тип C – N18, R72, S273 и R324;
тип D – K18, K72, C273 и S324-G325-K326;
тип E – K18, K72, C273 и R324,
где N – аспарагин, R – аргинин, C цистеин, S – серин, G – глицин, K – лизин.
Мы показали что, штаммы Y. pestis различных биоваров подвида microtus могут обладать одинаковыми типами LcrV, и различные типы LcrV могут быть свойственны разным популяциям Y. pestis одного биовара (рис. 1). Наиболее важным результатом нашего исследования является установление того факта, что LcrV типа D, характерный для эпидемически значимых штаммов Y. pestis subsp. pestis, также широко распространен среди штаммов различных биоваров subsp. microtus. 61 % штаммов подвида microtus обладали V антигеном этого типа.
Принимая во внимание тот факт, что V антиген D типа наиболее близок по аминокислотной последовательности к аналогичному белку своего "прародителя", Y. pseudotuberculosis [номер доступа в Gene Bank АDC 55500.1], а общее отличие от классического типа D для типов E и C замена трех C-концевых аминокислот (S324-G325-K326) на одну – (R324), которая является результатом вызванной двумя прямыми повторами (ATGACACG) делеции 3'-конца гена lcrV протяженностью 16 пар нуклеотидных оснований (Zhou et al., 2004) можно предположить, что наиболее молодой ветвью биовара caucasica является филогенетическая группа, общая для Присеванского горного (очаг 05), Зангезуро-Карабахского горного (очаг 06) и среднегорной степной части Приараксинского низкогорного (очаг 07) природных очагов чумы (LcrV типов E и C), а наиболее древней популяция штаммов, циркулирующих в Восточно-Кавказском высокогорном очаге (39) и дивергировавшая по данным MLVA25- типирования1 от клональных кластеров cc4 и cc5-7 до момента разделения последних (LcrV типа D) (рис. 2).
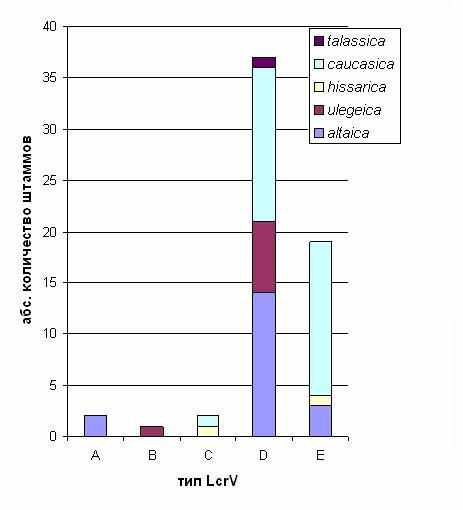
Рисунок 1 - Типы LcrV, характерные для штаммов Y.pestis subsp. microtus, принадлежащих к различным биоварам.
С учетом клональной теории возникновения и распространения возбудителя чумы в течение трех главных пандемий (Achtman et al., 1999, 2004; Devignat, 1951; Li et al., 2009), можно сделать вывод о том, что первый представитель вида Y. pestis обладал типом D аминокислотной последовательности LcrV, сохранившийся у потомков наиболее древних ветвей «полевочьих» штаммов и штаммов основного подвида. С другой стороны LcrV из штаммов Y. pestis, циркулирующих в популяциях разных видов полевок (Microtus spp.), проявляет все многообразие последовательностей (типы A–E). Это свидетельствует о том, что в организмах этого рода животных подобные мутации гена lcrV являются нейтральными или даже позволяют лучше адаптироваться к новой экологической нише.
Анализ нуклеотидных последовательностей гена lcrV и аминокислотных последовательностей V антигена послужили основой для компьютерного моделирования трехмерных структур выявленных вариантов белка LcrV2.
Трехмерную модель LcrV для "консенсусного" типа A рассчитали на основе кристаллической структуры LcrV типа D из штамма KIM (номер доступа 1R6F в базе данных SCOP). В соответствии со структурной классификацией белков (SCOP; версия 1.71) V антиген относится к классу мультидоменных протеинов, укладка белка LcrV содержит "α" домен, образованный преимущественно N-концевым участком, и "α+β" домен. Домены соединены антипараллельно скрученным витком.
Наибольший аминокислотный полиморфизм LcrV, включая замены в не моделированных участках белка, обнаружен в составе N-концевого участка (а.о. 1-146). На трехмерной молекулярной модели LcrV видно, что, за исключением а.о. 273, остальные замены (а.о. 72, 84, 103, 113 и 324) локализованы в составе "α" домена (рис. 3).
Мы попытались спрогнозировать влияние замены аминокислотных остатков LcrV из разных штаммов на значимые изменения трехмерной структуры белка. Кристаллическая структура LcrV из штамма Y. pestis KIM лишена целого ряда участков, невидимых на картах электронной плотности (Derewenda et al., 2004), поэтому мы создали более полную модель с использованием метода LGA.
Всего проведен структурный анализ шести вариантов аминокислотных замен в LcrV. Все замены, за исключением одной, выявлены либо в "гибких" участках молекулы, либо на поверхности белка и не приводят к изменению локальных химических свойств (гидрофобности, гидрофильности, амфипатичности, заряда) и, вероятно, оказывают минимальное влияние на структуру белка. Напротив, замена триптофана в позиции 113 на глютаминовую кислоту либо глицин должна оказывать значительное влияние на региональную структуру белка, а возможно и функции этого белка. Триптофан характеризуется большими размерами и, в основном, гидрофобностью, хотя и может образовывать одну водородную связь за счет азота в положении ε. Триптофан 113 в LcrV практически полностью скрыт в большом гидрофобном участке около центра N-концевого домена. Азот в положении ε стабилизирует структуру локальной петли водородной связью с карбонилом аспарагина 110 (рис. 4.5, A).
При замене триптофана глютаминовой кислотой, гидрофобные взаимодействия практически полностью утрачиваются, хотя глутамат и амфипатичен, и E113 более не может образовывать водородную связь с карбонилом N110. E113 может образовывать водородную связь с N144, который, в свою очередь, может образовывать водородную связь с карбонилом N110 для стабилизации локальной структуры петли. При замене W113 глицином (G113) утрачиваются все гидрофобные и водородные связи, что, вероятно, дестабилизирует петлю 108-112 и ведет к нарушению ее структуры, а, возможно, и к меньшей стабильности структуры всего белка (рис. 4, C). Действительно, в процессе выделения и очистки различных типов рекомбинантных LcrV было установлено, что антиген из штамма I-3455 (замена в положении 113 триптофана на глицин) существенно отличался от антигенов из других штаммов.
Суммируя все вышеизложенное можно констатировать, что белки LcrV из штаммов - представителей разных подвидовых групп Y. pestis обладают полиморфизмом размера, последовательности и трехмерной структуры.
Одним из основных факторов патогенности рода Yersinia является комплекс продуктов, кодируемых плазмидой pCad, присутствие которых необходимо для проявления вирулентности возбудителя (Portnoy, Falkow, 1981; Кокушкин, 1983). Мы установили, что белки LcrV штаммов - представителей различных внутривидовых групп Y. pestis обладают полиморфизмом аминокислотной последовательности, размера и трехмерной структуры. Можно предположить, что данный полиморфизм оказывает влияние на вирулентность штаммов с различными вариантами LcrV.

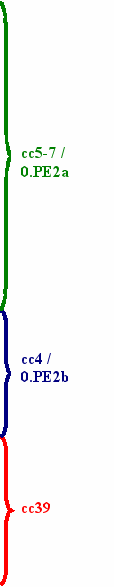
Рисунок 2 - Распределение типов аминокислотных последовательностей V антигена штаммов Y. pestis subsp. caucasica по кластерам NJ (Neighbor-Joining) дендрограммы MLVA25-типов исследованных штаммов. "Strain" – название штамма; "@" – штамм генотипирован in silico; "Isolation date" – время выделения; "F" – номер природного очага; "ssp" – подвид (P – pestis, MIC – microtus); "bv"- биовар (С – caucasica); MLVA-gt" – MLVA25-тип (Платонов, 2011); "AspA363" – аминокислота в позиции 363 аспартазы; "V-ag type" – тип V антигена.
Рисунок 3 - Модель структуры LcrV из штамма Y. pestis Angola представлена в виде стереоленточной диаграммы. Различия между нашей моделью и опубликованной ранее кристаллической структурой LcrV из штамма KIM выделены красным цветом. Места точечных различий между восемью использованными в исследовании штаммами представлены с боковыми цепями для каждого а.о. в виде шариков и палочек.
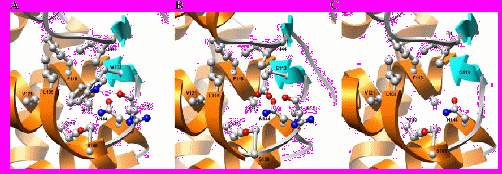
Рисунок 4 - Структуры участков трех типов LcrV Y. pestis, примыкающих к аминокислотному остатку 113. "Консенсусная" последовательность белка LcrV содержит в этом положении триптофан (A), а штаммы I-2359 и I-3455 – глютаминовую кислоту (B) или глицин (C), соответственно.
В качестве модели для сравнительной оценки вирулентности “полевочьих” штаммов, продуцирующих различные типы LcrV, использовали мышей и морских свинок (табл. 1).
Все изученные нами штаммы оказались высоковирулентными для белых мышей, что согласуется с данными других исследователей (Anisimov et al., 2004; Zhou et al., 2004b). При этом независимо от типа синтезируемого LcrV штаммы Y. pestis subsp. microtus были авирулентными для морских свинок (LD50 > 106 КОЕ).
Таким образом, выявленный нами структурный полиморфизм типов LcrV, секретируемых штаммами Y. pestis subsp. microtus, не влияет на их вирулентность для лабораторных животных, и "избирательная" вирулентность “полевочьих” штаммов обусловлена другими факторами.
Конструирование продуцентов V антигена Y.pestis
Работы по конструированию продуцентов V антигена для использования в составе диагностических и профилактических препаратов проводили с учетом полученных данных компьютерного моделирования трехмерной структуры LcrV и возможного влияния замен по отдельным аминокислотам на физико-химические, иммунохимические и протективные свойства белка. Мы клонировали ген lcrV из штаммов Y. pestis subsp. microtus биовара altaica I-3455 и I-2359, caucasica C-582, hissarica А-1728 и штамма Y. pestis subsp. pestis EV НИИЭГ в составе векторной плазмиды pET32b(+) в клетках протеазодефицитного штамма E. coli BL21(DE3) (Novagen, США). Рекомбинантные белки LcrV представляют собой пептиды с расчетной молекулярной массой 37,2 кДа.
Таблица 1 - Вирулентность “полевочьих” штаммов Y. pestis, продуцирующих различные структурные варианты LcrV
| Штамм Y. pestis | Тип LcrV | LD50 для лабораторных животных при п/к заражении (КОЕ) | |
| мыши | морские свинки | ||
| subsp. pestis | |||
| I-1996 | D | 1 (0-5)** | 8 (2-32) |
| 231 | D | 2 (1-8) | 4 (3-16) |
| subsp. microtus | |||
| bv. caucasica | |||
| C-535 | D | 12 (3-50) | 109 |
| С-376 | D | 3 (1-13) | > 106 |
| С-585 | Е | 6 (2-26) | > 106 |
| bv. hissarica | |||
| I-1249 | E | 32 (8-158) | > 106 |
| bv. ulegeica | |||
| I-2422 | B | 3 (1-13) | 109 |
| I-2239* | D | 10 | > 106 |
| I-3189* | D | 102 | > 109 |
| I-3190* | D | 102 | > 108 |
| bv. altaica | |||
| I-2359 | A | 50 (13-200) | > 108 |
| I-3259* | D | 102 | > 109 |
| * LD50 согласно паспортным данным. ** В скобках указаны доверительные интервалы для вероятности 95 . | |||
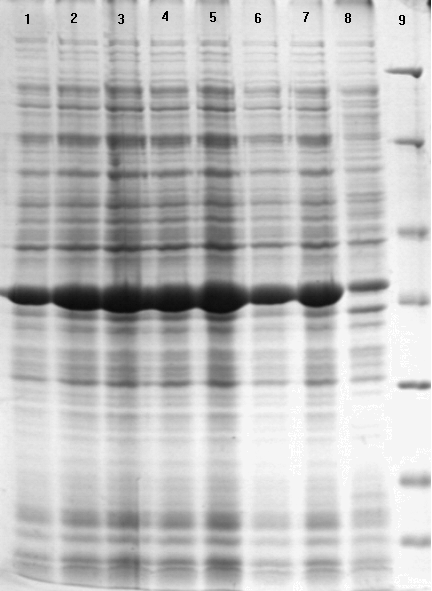
Аналитическая индукция синтеза V антигена (рис. 6) клетками E. coli BL21 (DE3), содержащими рекомбинантные плазмиды, показала присутствие рекомбинантного белка в виде мажорной полосы в экстракте индуцированных клеток, отсутствующей в экстракте клеток не индуцированных ИПТГ. Максимальную экспрессию V антигена наблюдали после трех часов индукции 1 мМ ИПТГ, причем целевой белок составлял не менее 50 % всех клеточных белков штаммов - продуцентов.
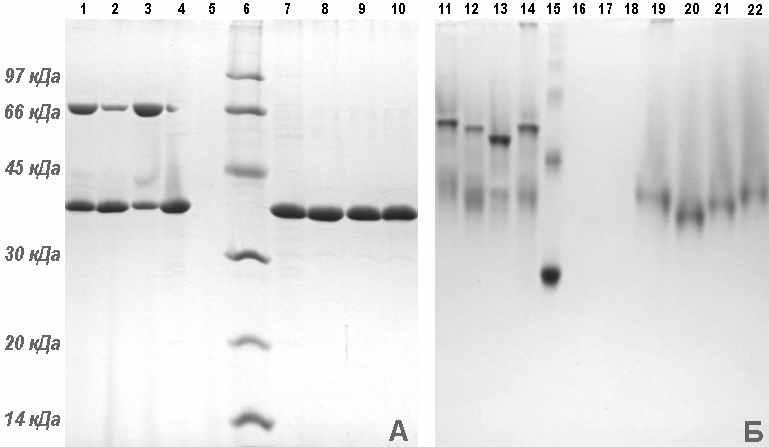
Выделение и хроматографическую очистку V антигена проводили в три этапа. На первом этапе V антиген очищали с помощью гель-эксклюзивной хроматографии, на втором этапе с помощью ионообменной хроматографии, на третьем этапе проводили гидрофобную хроматографию. Затем полученный препарат высокоочищенного V антигена подвергали диализу. Выход V антигена с чистотой не менее 95 % по результатам ДСН-электрофореза в редуцирующих условиях (рис. 7) составлял 35 мг/л.
Специфичность очищенного V антигена, выделенного из рекомбинантных штаммов E. coli, подтвердили в вестерн-блот анализе. В вестерн-блоте моноклональные антитела 2C3.3C7 и 5G5.E93 реагировали с белком LcrV в регионе, соответствующем 37 кДа (рис. 8). Несмотря на выявленные различия в структуре LcrV из различных штаммов Y. pestis оба моноклона обнаруживали все изучаемые варианты LcrV. В то же время они не реагировали с клеточным лизатом E. coli BL21(DE3).
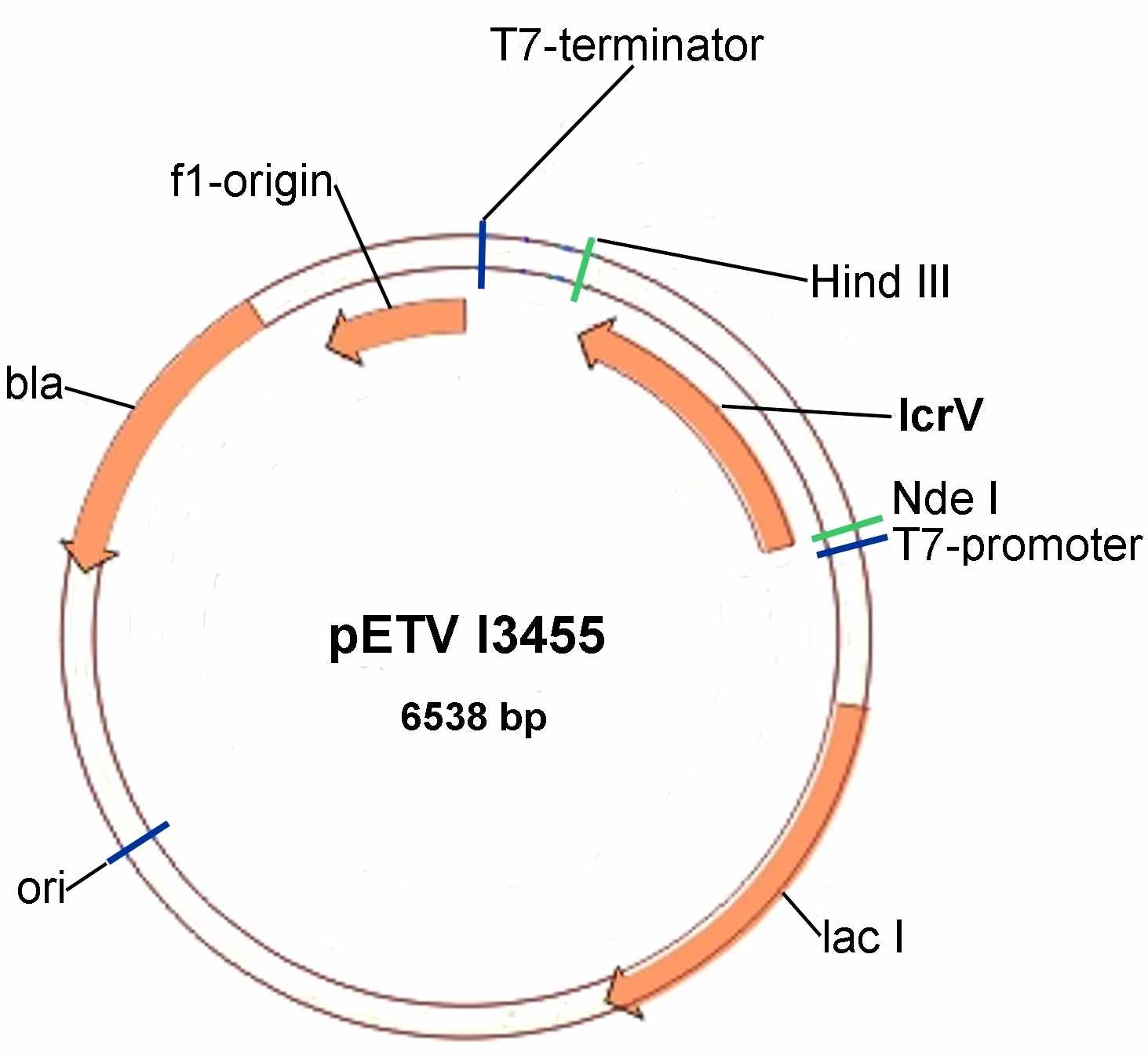
Рисунок 5 - Генетическая карта рекомбинантной плазмиды pETV I3455 (плазмиды pETV I2359, pETV C582, pETV А1728 и pETV9 имеют аналогичную структуру). T7-promoter - промотор бактериофага Т7; T7-terminator - терминатор бактериофага Т7; ori - точка начала репликации векторной плазмиды; bla - ген β-лактамазы (ApR); lacI – ген репрессора lac-оперона.
| А | Б | В |
 |  |  |
| | | |
Рисунок 6 - ДСН-ПААГ электрофорез рекомбинантных белков LcrV после индукции (окраска Кумасси голубым). (A) Линии: 1 - маркеры молекулярной массы (116,0, 66,2, 45,0, 35,0, 25,0, 18,4 и 14,4 кДа); 2-8 - клеточный лизат E. coli BL21(DE3)/pETV I2359 (2-7 после и 8 до индукции ИПТГ); 9-14 - клеточный лизат E. coli BL21(DE3)/pETV I3455 после индукции ИПТГ. (Б) Линии: 1 - маркеры молекулярной массы; 2-8 - клеточный лизат E. coli BL21(DE3)/ pETV C582 (2-7 после и 8 до индукции ИПТГ). (В) Линии: 1-8 - клеточный лизат E. coli BL21(DE3)/pETV A1728 (1-7 после и 8 до индукции ИПТГ); 9 - маркеры молекулярной массы (116,0, 66,2, 45,0, 35,0, 25,0, 18,4 и 14,4 кДа).
При изучении некоторых физико-химических свойств антигена мы показали, что прогревание белка LcrV при температуре 45 °С в течение 20 ч способствовало образованию олигомерных форм (рис. 9). Обработка проб перед электрофорезом 2-меркаптоэтанолом приводила к исчезновению высокомолекулярных полос во всех соответствующих пробах как на нативном геле (рис. 9 Б), так и на геле с додецилсульфатом натрия (рис. 9 А). Таким образом, стабильность агломерированного V антигена поддерживалась дисульфидными связями.
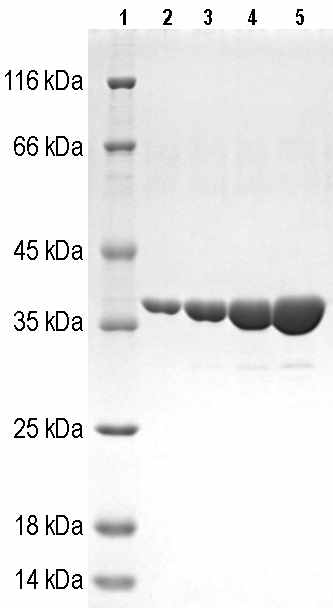
Рисунок 7 - ДСН-ПААГ электрофоретический анализ LcrV из штамма E. coli BL21(DE3)/pETV I3455 после хроматографической очистки: Линия 1 маркер молекулярных масс, линии 2 , 3, 4, 5 0,5 мкг, 1 мкг, 2 мкг, 5 мкг белка LcrV соответственно.
Рисунок 8 - ДСН-ПААГ и соответствующий иммуноблот с МкАТ к LcrV 2C3.3C7 и 5G5.E9. Линии: 1 - клеточный лизат E. coli BL21(DE3)/pETV A1728 после индукции; 2 - клеточный лизат E. coli BL21(DE3)/pETV С582 после индукции; 3 - клеточный лизат E. coli BL21(DE3)/pETV I2359 после индукции; 4 - клеточный лизат E. coli BL21(DE3)/pETV I3455 после индукции; 5 – клеточный лизат E. coli BL21(DE3)/pETV 9 после индукции; 6 – клеточный лизат E. coli BL21(DE3) (отрицательный контроль).