
Ослин навчальний посібник до лабораторних занять з фізіології рослин для студентів біологічного факультету Видавництво поліграфічний центр "Київський університет" 2004
| Вид материала | Документы |
- Мусієнко М. М., Панюта О. О. Біотехнологія рослин. Навчальний посібник, 5445.72kb.
- Ділова іноземна мова" для студентів геологічного факультету (магістрів V-VI курсів), 329.16kb.
- Савицький В. М., Хільчевський В. К., Чунарьов О. В., Яцюк, 2100.06kb.
- Вченої ради біологічного факультету протокол №11 від 25 квітня 2005, 115.21kb.
- Тематичний план занять «Патофізіологія ендокринної та нервової систем» Для студентів, 1055.52kb.
- Київський університет туризму, економіки І права «Основи готельного менеджменту», 5077.65kb.
- Полтавський національний педагогічний університет імені В. Г. Короленка Історичний, 675.02kb.
- Практикум з педагогіки вищої школи: Навчальний посібник за модульно-рейтинговою системою, 411.72kb.
- Навчальний посібник для студентів філологічних спеціальностей вищих педагогічних навчальних, 3345.03kb.
- Практичний курс англійської мови навчальний посібник з практики усного та письмового, 5352.9kb.
Робота № 9. Вимірювання відстаней та прокладання профілів
та трансект на воді.
Теоретичні передумови. Відстані на водоймі вимірюють за допомогою мірного шнура. Мірний шнур виготовляють із різних синтетичних і натуральних мотузків або крученого пенькового шпагату. Шнур із пенькового шпагату, який правильно виготовлений ( вимочений, розтягнутий і висушений ) під час роботи на воді не плутається, не розтягується і позначки на ньому як в сухому, так і в вологому стані мають постійні відстані.
Шнурок зручно розмітити ганчірочками або прапорцями чорного, білого, синього і червоного кольорів. Чорними позначають непарні поділки, білими – парні, синіми – кожну п`яту, червоними – кожну десяту позначки.
Мірний шнур повинен бути довжиною близько 250- 300 м і складатися із кількох відрізків по 40-50 м кожний, які на кінцях мають карабіни для зчеплення.
При розтяганні шнура на водоймі під час прокладання профілів до нього через кожні 20 м прикріплюють поплавки із пінопласту чи іншого легкого матеріалу, або дерев`яні дощечки пофарбовані білою або червоною олійною фарбою. Поплавки можна не відчіпляти від шнура впродовж всієї роботи, намотуючи 40 або 50-метрові куски шнура на ці ж поплавки.
На маленьких водоймах (ставках, невеликих озерах, річках ) шнур легко розтягається з одного берега на інший і великих труднощів не викликає. На великих водоймах на мілководді, яке заростає, шнур розтягають трохи далі межі розповсюдження рослин. Шнур краще розтягати в напрямку з води на берег.
Мета роботи – навчитися вимірювати відстані та прокладати профілі і трансекти на водних об’єктах.
Матеріали та обладнання: човен, поплавок з вантажем, мірний шнур, журнал для записів.
Хід роботи. На середині водойми, за кілька метрів до межі розповсюдження водних рослин встановлюють поплавок з вантажем до якого причіплюють мірний шнур. На човні повільно рухаються з середини водойми до берега розмотуючи шнур і постійно вирівнюючи поплавки за його допомогою. На березі шнур міцно прив’язують. Мірним шнуром вимірюють протяжність заростей водних рослин за горизонтальним зональним розподілом. Опис профілів починають з короткої характеристики прибережної рослинності, потім описують водну рослинність, виділяючи в її межах окремі фітоценози. Кількість профілів, які слід прокласти в тій чи іншій водоймі для опису, кількісного обліку і картування рослинності, залежить, насамперед, від характеру заростання водойми та її величини. В маленьких, мілководних водоймах з незначним різноманіттям біотопів і рослинності, можна прокласти 1-2 профілі. Після прокладання профілів описують та обліковують рослинність за профілями. Замальовують профілі ділянки водойми з водними рослинами. Роблять висновки.
Робота № 10. Проведення картування водної рослинності
на водоймі.
Теоретичні передумови. Перед тим, як детально описувати та проводити картування рослинності на водоймі, слід провести рекогносціювальний об’їзд водойми (або її частини) на човні або обхід її з берега, для того, щоб ознайомитися з характером рослинності і загальними рисами розподілу її угруповань. Під час первинного аналізу відмічають характерні ділянки водойми, на яких буде проведено детальніші дослідження.
Для того, щоб мати змогу показати хоча б приблизно розподіл рослинності на водоймі і визначити площі, що займають окремі рослинні угруповання, потрібні великомасштабні карти, бажано такі, на яких були б нанесені і глибини. На картах далеко не завжди вдається відтворити дійсну картину розподілу окремих одиниць рослинності і визначити в подальшому площі, які вони займають. Тому, крім загальної картосхеми для деяких найбільш цікавих, щільно зарослих ділянок водойми, які зумовлюють інтерес в дослідників, складають плани великого масштабу ( навіть до 10 м в 1 см). На ці плани вже можна нанести і фрагменти фітоценозів, які займають зовсім незначні ділянки. При складанні картосхем і планів не можна обмежуватися нанесенням меж лише видимої рослинності. Слід також нанести межі розповсюдження не видимих зверху угруповань, занурених та невеликих придонних рослин, для виявлення яких треба користуватися відповідними приладами.
Складати картосхеми розподілу рослинності можна візуально з човна, вимірюючи відстані та протяжність різних типів рослинності гребками, мірним шнуром, або рулеткою (на березі).
Для оформлення картосхеми розподілу рослинності у водоймі для позначення різноманітних одиниць рослинності (асоціацій, формацій та ін., залежно від завдання дослідженнь), які наносять на карту, користуються умовними позначками – різними типами штриховок або знаками. Види рослин на картах, якщо це потрібно, можна позначити початковими буквами їхньої родової або видової назви.
Позначаючи штриховкою формацію надводних рослин використовують штриховку з нахилом вправо, плаваючих – хвилясту горизонтальну лінію або косу клітинку, занурених – вертикальну штриховку. Штриховки повинні бути рідкими, щоб надати можливість вписати в них букви, якими позначають види рослин.
Щоб провести глазомірну оцінку заростання водойми використовують схему для визначення ступеня заростання з наступними позначеннями:
+5 – заростання надмірне, рослинністю вкрито понад 50% поверхні водойми,
5 – дуже велике ( від 1/3 до 1/2 поверхні, 36-50 %),
4 – велике (від 1/5 до 1/3 поверхні, 21- 35%),
3 - середнє (від 1/10 до 1/5 поверхні, 11-20%),
2 – невелике (від 1/50 до 1/10 поверхні, 3 –10%),
1 – незначне (від 1/100 до 1/50 поверхні, 1-2%).
За картами визначають площу, яка зайнята рослинністю у водоймі, а також площу її окремих угруповань.
Мета роботи – провести картування розподілу водної рослинності на природній водоймі, скласти картосхеми водойм та плани великого масштабу з фітоценозами водних рослин.
Матеріали і обладнання: карта (або схема) досліджуваної водойми, визначник рослин, журнал для записів, мірний шнур або рулетка.
Хід роботи. На природній водоймі окомірно провести оцінку заростання поверхні водойми та берегової смуги, визначити площу заростання у відсотках та балах. Створити картосхему дослідженої водойми та скласти плани великого масштабу найхарактерніших ділянок водойми, нанести на них фрагменти фітоценозів, позначивши штрихуванням різні екологічні групи рослин та умовними позначками (або літерами) окремі види водних рослин. За допомогою створених картосхем та великомасштабних планів визначити площу окремих угруповань водних рослин. Зробити висновки.
Лабораторні методи дослідження водних рослин.
Анатомічні методи дослідження водних рослин.
Робота № 11 . Особливості анатомічної будови водних рослин
різних екологічних груп.
Теоретичні передумови. Відмінності фізико-хімічних властивостей середовища, в якому відбувається ріст і розвиток вищих водних рослин (води, ґрунту, повітря) створюють для них різкоконтрастні умови кисневого, світлового, температурного режимів, забезпечення елементами вуглецевого та мінерального живлення. Водне середовище, порівняно з атмосферою містить недостатньо вуглекислого газу і кисню, більшу густину та інший температурний режим. Фотосинтез водних рослин відбувається за невисокої інтенсивності світла та іншого спектрального складу у занурених у воду частинах рослин. Поверхневі шари води поглинають червоні промені і на значну глибину доходять лише сині.
Все це зумовлює виникнення у вищих гідромакрофітів цілого ряду зовнішніх і внутрішніх адаптацій, які супроводжуються відповідними змінами метаболізму і анатомо-морфологічної будови. Із збільшенням гідрофільності рослин, тобто в напрямку від групи повітряно-водних (гелофітів) до власно водних (гідатофітів), в рослинних тканинах збільшується частина повітряних каналів – аеренхіми, поступово відбувається редукція покривних, механічних тканин та провідної системи рослин, спрощується будова фотоасимілюючих органів, кореневої системи, інколи до повного її зникнення.
Деякі вищі гідромакрофіти повністю втрачають функціонально відокремлені вегетативні органи (листок, стебло, корінь), набуваючи вигляду так званих листеців, які мають дуже спрощену анатомо-морфологічну будову, але виконують всі фізіологічні функції втрачених органів. Перераховані зміни будови водних рослин спостерігаються не лише у різних видів, що ростуть в певних умовах (на березі, мілководді, або на значній глибині), а й у одного виду і, навіть, в різних органах однієї рослини, залежно від глибини місцезростання цієї рослини, тобто фактори довкілля виконують роль індукторів тих або інших змін в їх будові. Зовнішні і внутрішні адаптації водних рослин до умов життя у водоймі супроводжуються відповідними перебудовами і на рівні метаболізму.
Мета роботи – провести порівняльний аналіз анатомічної будови водних рослин різних екологічних груп.
Матеріали та обладнання: живі та фіксовані органи водних рослин різних екологічних груп (стебла, листки, корені, кореневища, столони); світловий мікроскоп, предметні і накривні скельця, препарувальні голки, леза; флороглюцин, соляна кислота.
Хід роботи. Роблять поперечний зріз органу рослини лезом безпечної бритви, обробляють зріз флороглюцином і соляною кислотою, розглядають його будову під світловим мікроскопом, відмічають особливості будови, які характерні для певної екологічної групи рослин і зарисовують схематично будову кожного органа.
Анатомічна будова органів очерету звичайного, як представника гелофітів.
Стебло. У очерета звичайного на поперечному розрізі можна побачити диференціацію стебла: шкірочку, кору і осьовий циліндр (рис.9).
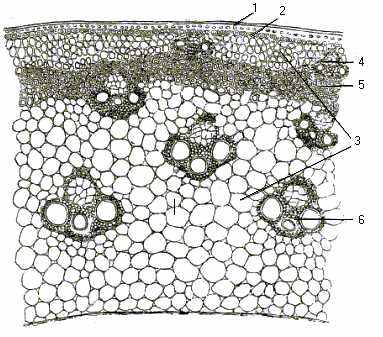
Рис. 9. Стебло очерету звичайного (поперечний розріз, збільшення 80): 1- епідерма; 2- підепідермальний шар; 3- паренхіма; 4- кора; 5- склеренхімне кільце; 6- провідні пучки.
Під шкірочкою знаходиться первинна кора, яка складається з кількох шарів хлорофілоносних паренхімних клітин; внутрішній шар клітин представлений ендодермою. Осьовий циліндр має багатошаровий перицикл у вигляді склеренхімного кільця. На поперечних розрізах простежуються численні пучки, які розташовані рядами. У стеблі молодого очерету чітко видно три кільця досить великих пучків. У ксилемній частині пучків видно дві великі пористі судини, які розташовані симетрично. Між пучками лежать 1-2 великі судини. Ксилема розташована підковоподібно навколо флоеми, яка складається із правильно розташованих сітчастих трубок. Пучок має механічну обкладку – склеренхімну піхву. Арматура стебла очерету звичайного складається із товстостінної шкірочки, склеренхімного кільця і механічних обкладинок провідних пучків. Основна тканина в центральній частині стебла складається з великих тонкостінних клітин. Частина основної паренхіми руйнується і в дорослому стані стебло очерету звичайного стає порожнім, за винятком вузлів. У очерету звичайного в стеблі дорослої рослини в коровому шарі утворюються повітряні порожнини.
Кореневище. В кореневищі очерету раніше, ніж в надземних пагонах, набуває розвитку система покривних тканин у вигляді шарів клітин відносно товстої первинної кори ( рис.10).
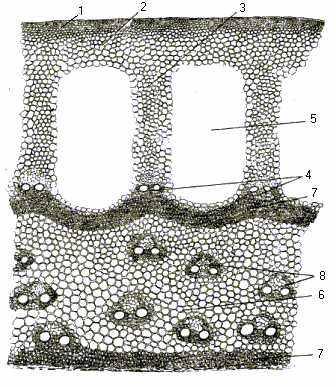
Рис. 10. Багаторічне кореневище очерету звичайного (поперечний розріз, збільшення 48): 1- епідерма; 2- хлорофілоносна паренхіма; 3- тяжі; 4- кора; 5- повітряні порожнини; 6- центральна порожнина; 7- склеренхімне кільце; 8- провідні пучки.
Епідермис кореневища складається з коротких та довгих клітин з хвилястими стінками, що чергуються. В кореневищі чітко виражені ділянки первинної кори та осьового циліндру. Первинна кора має великі повітряні порожнини, які розділені між собою перегородками – тяжами. З віком ці тяжі збільшуються за рахунок збільшення кількості клітин, що їх складають. Потовщення оболонок клітин, які утворюють тяжі, збільшує пружність та стійкість кореневища.
Механічна система тканин сильно розвинена. Арматура кореневища у очерету звичайного представлена переферичним склеренхімним кільцем і механічними обкладинками провідних пучків. З віком відбувається потовщення прикорового склеренхімного кільця, а також утворюється друге склеренхімне кільце, що оточує внутрішню порожнину кореневища.
Провідні пучки розкидані в центральній частині, а в місцях закінчення тяжів розташовані так звані корові пучки.
Центральна частина кореневища, порівняно з коровою, розвинена особливо сильно. Це зумовлено тим, що вона виконує функцію вмістилища запасних речовин. На відміну від столонів, кореневища мають потовщені стебла і довше залишаються життєздатними.
Столони. На столонах розвиваються не лише бруньки, а й корені. Столони інколи поширюються на десятки метрів і розвиваються наземно. Для них характерний розвиток подовжених міжвузлів і мала товщина стебла. За анатомічною будовою столони подібні до стебел, але несуть деякі органи, які характерні для кореневищ, - додаткові корені.
Корінь. Кора кореня очерету звичайного складається із зовнішньої частини – екзодерми, внутрішньої частини – ендодерми і, розташованих між ними, паренхімних тонкостінних клітин з целюлозними стінками. В зовнішній частині кори ці клітини багатогранні і щільно зімкнені. В паренхімному шарі клітини округлі і розташовані правильними радіальними рядами. Між клітинами паренхіми, в результаті розщеплення клітинних стінок, утворюються великі міжклітинні повітряні канали. Ендодерма складається із щільно зімкнених паренхімних клітин з целюлозними стінками, що мають на поперечному розрізі форму прямокутників із закругленими кутами.
Всередину від ендодерми в корені розташований осьовий циліндр. Переферична його частина (перицикл) - це кільце живих паренхімних, потовщених та здерев’янілих клітин. Далі від перициклу розташована провідна система; це складний радіальний пучок, що складається із ділянок ксилеми і флоеми, які чергуються. Судини ксилеми розкидані між основною тканиною. З віком анатомічна будова кореня очерету залишається майже незмінною, за винятком склерифікації – потовщення та здерев’яніння оболонок клітин.
Кореневі волоски утворюються біля верхівки кореня з клітин епіблеми. Після відмирання кореневих волосків клітини епіблеми, які розташовані під волосками, також відмирають. В зоні кореня, де відмирає епіблема, в екзодермі відбуваються деякі зміни і вона стає захисним шаром.
Листок. Для лінійних листків очерету звичайного, як і взагалі для всіх злакових, характерний інтеркалярний ріст, тобто в базальній частині їх зберігається зона меристематичних клітин, поділ яких зумовлює витягування листка. Ззовні листок вкриває шкірочка – епідерміс (рис.11).
Одношарова кутикула верхнього епідермісу листка очерету, як і нижнього, порівняно товста. Продихи розташовані з обох боків листка, але з нижнього боку їх більше, ніж з верхнього. Очерет має злаковий тип продихового апарату, особливість якого полягає в тому, що клітини-замикачі продихів мають вигляд вузьких прямокутників гантелеподібної форми. Клітини шкірочки розташовані правильними, майже паралельними рядами з хвилястими поздовжніми стінками, причому в ряду чергуються довгі й короткі стінки.
У очерету звичайного в шкірочці і в верхніх шарах мезофілу розташовані пухирцеподібні (водоносні) клітини, які відіграють важливу роль під час згортання та розгортання листка. Ці клітини сильно відрізняються від клітин епідермісу не лише тонкими стінками і великими розмірами, а й тим, що в них клітинна порожнина сильно розширюється в напрямку від поверхні листка до його середини. Водоносні клітини розташовані долинці між ребрами листкової пластинки.
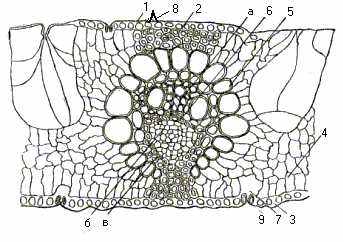
Рис. 11. Листок очерету звичайного (поперечний розріз, збільшення 185):
1- кутикула; 2- верхня епідерма; 3- нижня епідерма; 4- паренхіма; 5- клітини обкладки пучка; 6- провідний пучок (а- ксилема, б- флоема, в- механічна тканина); 7- повітряна порожнина; 8- шипик; 9- продихова щілина.
Великі провідні пучки проходять уздовж листка майже паралельно, з’єднуючись між собою перемичками. Елементи флоеми і ксилеми, які розташовані в центральній частині пучка оточені облямівкою з щільно зімкнених видовжених великих клітин, що являють собою систему активно асимілюючого апарата. У провідному пучку розташований склеренхімний тяж, який закінчується під шкірочкою. Значний шар склеренхімних клітин забезпечує механічну міцність листка. В листку очерету немає типових палісадної і губчастої хлоренхім, асиміляційна тканина складається з гіллястих паренхімних клітин.
Анатомічна будова органів глечиків жовтих, як представника
вкорінених плейстофітів.
Кореневище. Стебло глечиків жовтих перетворено на товсте кореневище, яке розташоване горизонтально в донних покладах. Кореневище – це видозмінене стебло, що виконує функцію органа, в якому відкладаються запасні поживні речовини. Судин в кореневищах немає, а ксилема має кільчасті або спіральні трахеїди. В стеблах і кореневищах гарно розвинена система повітряних каналів. Механічні тканини розвинені гірше, ніж у гелофітів.
Листок. Водний спосіб життя цієї групи рослин, який постійно пов’язаний з дефіцитом світла і кисню, спричинив гетерофілію – утворення у багатьох з них листків двох типів – плаваючих і занурених. Мезофіл плаваючого листка з обох боків вкритий одношаровою епідермою (рис.12).
На верхньому боці її добре розвинена кутикула з восковим нальотом, завдяки чому поверхня листка не змочується і газообмін не порушується. Продихи є лише на верхній частині листка, вони завжди відкриті. У глечиків їх близько 400 на 1 мм2 листкової пластинки. У плаваючому на воді листку не буває водного дефіциту, а тому й немає потреби регулювати транспірацію. Навпаки, постійне випаровування води захищає листок від перегрівання. На нижньому боці його можна побачити зкорковілі клітини – гідропоти, які, очевидно, сприяють кращому газообміну.
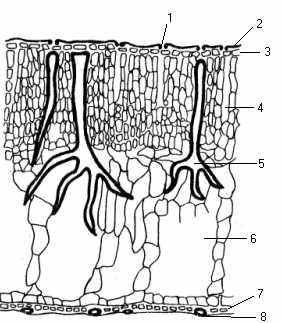
Рис. 12. Листок глечиків жовтих (поперечний розріз, збільшення 280): 1 – продих; 2 – восковий шар; 3 – верхня епідерма; 4 – мезофіл; 5 – опорні клітини; 6– повітряна порожнина; 7 – гідропота; 8 – нижня епідерма.
Мезофіл диференційований на багатошарову палісадну тканину майже без міжклітинників і губчасту з великими повітряними порожнинами, які безпосередньо прилягають до нижньої епідерми. Система повітряних порожнин в плаваючих на воді листках забезпечує їх плавучість та інтенсивний газообмін із зануреними у водне середовище частинами рослин.
Занурені листки серцеподібно-стрілоподібні. Листкові пластинки дуже тонкі, не мають кутикули і вкриті одношаровою епідермою. Мезофіл однорідний, складається лише з двох шарів клітин з великими міжклітинниками. В мезофілі утворюються повітряні порожнини, які прилягають, безпосередньо, до епідерми. В цьому відношенні вони дещо нагадують тіньові листки мезофітів, у яких кількість рядів палісадної тканини, порівняно з світловими, зменшується. Відсутність палісадної тканини в зануреному листку пов’язана із зниженням інтенсивності світла в товщі води.
В листках слабко розвинені механічні й провідні тканини.