
 Авторефераты по всем темам >>
Авторефераты по биологии
Авторефераты по всем темам >>
Авторефераты по биологии
ВЫСШИЕ УРОВНИ ОРГАНИЗАЦИИ ХРОМТИНА И РЕГУЛЯЦИЯ ФУНКЦИОНАЛЬНОЙ АКТИВНОСТИ ГЕНОМА
Автореферат докторской диссертации по биологии
|
Страницы: | 1 | 2 | 3 | 4 | 5 | 6 | |
 СКАЧАТЬ ОРИГИНАЛ ДОКУМЕНТА
СКАЧАТЬ ОРИГИНАЛ ДОКУМЕНТА3.7.аа Роль конденсинов в структурной организации митотических хромосом.
Для анализа роли конденсинов в компактизации хромосом в митозе важно установить хронологию ассоциации архитектурных белков хромосом с конденнсирующимися профазными хромосомами и их пространственную локализацию. В клетках, находящихся в 02-фазе, топоизомераза II alpha распределяется в объеме ядра более или менее диффузно, за исключением нескольких ярких фонкусов, соответствующихся реплицированным центромерам (Rattner et al, 1996). В противоположность топоизомеразе II, SMC2 локализуется в виде мелких тончек, часто концентрирующихся на периферии масс конденсированного хроматинна. На этой стадии колокализации SMC2 и топо II практически не наблюдается.
В ранней профазе окрашивание SMC2 более отчетливо концентрируется в мелких фокусах, ассоциированных с периферией хромосом. На этой стадии конроткие параллельно лежащие линейные 8МС2-позитивные структуры начинают выявляться на поверхности хромосом. Топоизомераза II распределяется более
30
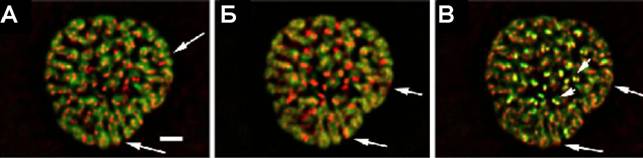
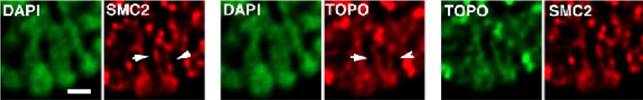
Рис.14. Распределение SMC2 и топоизомеразы IIв средней профазе. (А) ДАПИ (зеленый) и SMC2 (красный); (Б) ДАПИ (зеленый) и топоизомераза II(красный); (В) топоизомерза II(зеленый) и SMC2 (красный). Стрелки указывают на области, увеличенные на врезках. В средней профазае окраска SMC2 и топоизомеразы IIначинают частично совпадать, формируя колинеарные участки, расположенные преимущественно на поверхности хромосом (А-В, врезки, короткие стрелки).
диффузно, чем SMC2. Однако, в отличие от G2, в ранней профазе часто наблюндается колокализация 8МС2-позитивных фокусов с парными фокусами топонизомеразы II, соответствующим центромерам. Вне центромерных районов топо II и SMC2 практически не колокализуются.
В средней профазе колокализации SMC2 и топоизомеразы II становится еще более выраженной, но SMC2 в большей степени локализуется на перифенрии конденсированных участков хромосом. Области колокализации включают протяженные (до 1-2 мкм) линейные окрашенные структуры, которые преимунщественно располагаются на периферии хромосом. Интересно, что в некоторых случаях линейные сегменты топо II и SMC2 располагаются параллельно друг другу, но не колокализуются полностью, и только в центромерных районах ярнкие точки топо II и SMC2 полностью совпадают.
Только в поздней профазе колоклизация топо II и конденсинов становится очевидной во всех областях ядра. Топо II в основном концентрируется в центронмерах, но также четко видно окрашивание топо II и SMC2 вдоль плеч хромосом. При этом наблюдается как периферическая, так и осевая локализация этих окра-
31

Рис. 15. Аксиальная локализация SMC2 в метафазных хромосомах. Стереопары полутонких срезов изолированных метафазных хромосом, окрашенных Nanogold-конъюгатами антител с серебряным усилением, выявляют осевое распределение SMC2 в виде зоны шириной 0.15-0.2 мкм
шенных зон, а к началу метафазы осевая локализация топоизомеразы II и SMC2 становится очевидной. В полностью компактизованных хромосомах топо II и SMC2 демонстрируют высокую степень колокализации. Иммуноэлектронная микроскопия нативных метафазных хромосом позволила выявить внутри метанфазных хромосом зоны окрашивания антителами против SMC2 шириной 0.15-0.2 мкм, что составляет примерно одну треть от общего диаметра хроматиды (0.5 мкм).
Таким образом, в ходе митотической компактизации хромосом наблюданется корреляция между морфологическими изменениями хромосом и динаминческим перераспределением топо II и 8МС2-субъединицы конденсина. При этом первичное распределение топо и SMC в значительной степени связано с поверхностью компактизующихся хромосом, за исключением центромерных районов. Перераспределение этих белков с периферии в осевые области хронмосом происходят значительно позже, чем завершается формирование равнонмерно упакованных хромосом со сформированной осью, характерное для среднней профазы.
32
3.8. Исследование поведения конденсинов при искусственном изнменении степени компактизации хромосом в живых клетках.
окализация белков ХСАР-Е и pEg7 в митотических клетках при искусственной декомпактигации хромосом в условиях обратимого гипотонического шока.
Искусственную деконденсацию хромосом в живых клетках вызывали инкубацией клеток в гипотонической среде (30% среды L-15). Поскольку такое воздействие является структурно обратимым (см. выше), можно установить связь между присутствием конденсинов и структурной целостностью митотиченских хромосом.
Обработка живых клеток 30% раствором среды L-15 в течение 5 минут вызывает частичную деконденсацию митотических хромосом, общая морфология хромосом при этом сохраняется. Через 15 минут инкубации начинается постепенная адаптация клеток к гипотонической среде, сопровождающаяся реконденсацией хромосом; примерно через 30 минут внешний вид хромосом близок к исходному. При этом в фазе деконденсации наблюдается частичный выход субъединиц конденсина ХСАР-Е и pEg7 в цитоплазму. В то же время, в осевых областях деконденсированных хромосом сохраняется повышенная концентрация конденсинов. Таким образом, деконден-сация хромосом in vivo не коррелирует с диссоциацией конденсинов, хотя ненльзя исключить, что остающиеся связанными с хромосомами конденсиновые комплексы в этих условиях теряют активность и не выполняют своей функции стабилизации структуры хромосом. Через 30 минут после начала воздействия, когда наступает фаза реконденсации, антитела к pEg7 и ХСАР-Е перестают окрашивать хромосомы и выявляются преимущественно в цитоплазме. Однако диссоциация конденсинов не препятствует успешной реконденсации хромосом и восстановлению структуры хромонемных элементов. Таким образом, восстанновление компактной структуры хромосом после гипотонического шока происнходит без участия конденсинов, или по крайней мере, только незначительная доля популяции конденсинов, не выявляемая методами иммунофлуоресцентной микроскопии, остается связанной с митотическими хромосомами. Эти данные противоречат как общепринятым представлениям о конденсиновом комплексе как об интегральном компоненте хромосомного скэффолда, обеспечивающего специфическую радиально-петлевую организацию хроматина в интерфазных и митотических хромосомах (Gasser, 1995; Hirano, 1998), так и гипотезе о возможнной энзиматической роли конденсинов, участвующих в активном реконфигури-ровании топологически замкнутых петлевых доменов хроматина в ходе конденнсации хромосом (Kimura, Hirano, 1997; Kimura et al, 1999).
33
окализацияаа белковаа ХСАР-Еаа иаа pEg7аа ваа митотическихаа клеткахаа после воздействия цитостатиков.
Для стабилизации хромосом в живых клетках в компактном состоянии длительное время использовали цитостатики нокодазол и таксол. Обработка нокодазолом приводит к деполимеризации микротрубочек, и полному разрушению митотического веретена, таксол, напротив, понижает критическую концентрацию полимеризации тубулина, что приводит к формированию дополнительных нецентросомальных фокусов полимеризации микротрубочек в цитоплазме митотических клеток и вызывает блок митоза вследствие нарушения обмена тубулина. При обработке клеток 0.5 мкг/мл нокодазола или 10 мкг/мл таксола в течение 6 часов происходило накопление метафазных клеток, при этом хромосомы в них сохраняли компактную структуру. В данных условиях не наблюдалось реверсии митоза, то есть перехода клеток в интерфазу без расхождения хромосом. При этом в клетках, обработанных нокодазолом, антитела к pEg7 и к ХСАР-Е перестают окрашивать хромосомы уже через 3 часа после начала воздействия. В клетках, инкубированных в среде, содержащей таксол, также наблюдается постепенный выход белков pEg7 и ХСАР-Е из митотических хромосом. В качестве контроля использовались антинтела к топоизомеразе II, которая в метафазе локализуется вместе с конденсина-ми в осевых районах хромосом. Оказалось, что в этих условиях антитела успешно выявляют топоизомеразу II в осевых областях компактизованных хромосом, что делает маловероятным предположение об ограниченной диффунзии антител против субъединиц конденсинового комплекса внутрь компактизонванных хромосом.Данные наблюдения являются дополнительным аргументом против непосредственной структурной роли конденсинов в митотических хронмосомах. Возможно, конденсины необходимы только на начальных этапах формирования высших уровней компактизации ДНК в митотических хромосомах, а не для стабилизации их макромолекулярной структуры, которая достигается в результате действия других структурных белков. Постоянное присутствие конденсинов на хромосомах в течение всего митоза может обеспечивать их пассивное перераспределение в дочерние ядра.
3.9.а Конденсиновый комплекс в мейозе.
На стадии диплотены мейотической профазы I хромосомы амфибий принобретают конфигурацию ламповых щеток, характерную наличием осевых структур, содержащих высококонденсированный транскрипционно неактивный хроматин, и отходящих от него петель хроматина, содержащих активно транснкрибирующиеся гены. Хромосомные петли представлены сильно деконденсиро-ванными фибриллами хроматина, на которых размещены активно работающие РНК-полимеразы П. При этом, петли составляют лишь небольшую долю хронмосомной ДНК, в то время как основная масса ДНК присутствует в компактной
34
форме в осевых районах хромосом-гомологов, каждый из которых представлен двумя сестринскими хроматидами, сохраняющими тесную когезию. Осевые районы демонстрируют линейную неоднородность в содержании хроматина, выражающуюся в формировании многочисленных последовательно располонженных гранул высококомпактизованного хроматина, называемых хромомера-ми, чередующихся с участками относительно менее плотной упаковки хроматинна. Мы изучили внутриклеточную локализацию конденсинов в ооцитах Хепо-pus laevis на стадии профазы мейоза I, обращая особое внимание на их взаимондействие с профазными хромосомами типа ламповые щетки.
Для исследования субклеточного распределения ХСАР-Е и pEg7, микронхирургическим методом были получены цитоплазматическая и ядерная фракнции ооцитов и присутствие конденсинов в белковых экстрактах этих фракций было проанализировано методом Вестерн-блоттинга. Оба белка главным обнразом выявляются в цитоплазматической фракции ооцитов. Наиболее выраженнную цитоплазматическую локализацию демонстрирует XCAP-D2, присутствие
|
Страницы: | 1 | 2 | 3 | 4 | 5 | 6 | |
Авторефераты по всем темам >>
Авторефераты по биологии