
 Авторефераты по всем темам >>
Авторефераты по биологии
Авторефераты по всем темам >>
Авторефераты по биологии
ВЫСШИЕ УРОВНИ ОРГАНИЗАЦИИ ХРОМТИНА И РЕГУЛЯЦИЯ ФУНКЦИОНАЛЬНОЙ АКТИВНОСТИ ГЕНОМА
Автореферат докторской диссертации по биологии
|
Страницы: | 1 | 2 | 3 | 4 | 5 | 6 | |
 СКАЧАТЬ ОРИГИНАЛ ДОКУМЕНТА
СКАЧАТЬ ОРИГИНАЛ ДОКУМЕНТАНа распластанных препаратах митотических хромосом, приготовленных с использованием 15% раствора Хэнкса вместо стандартного 75 мМ КС1, мета-фазные хромосомы выглядят сильно деконденсированными и не обнаруживают признаков продольной или поперечной неоднородности. Аналогичная обранботка митотических хромосом, образовавшихся после реадаптации клеток к первому гипотоническому шоку, вызывает дифференциальную деконденсацию хромосом, в составе каждой хроматиды которых без дополнительных обрабонток выявляются отчетливые аксиальные глобулы, располагающиеся симметричнно в сестринских хроматидах. Рисунок расположения аксиальных гранул марнкерных хромосом выявил частичное соответствие этих структур G-бэндам, при этом наблюдалось 30%-ное удлинение хромосом по сравнению с контролем. По-видимому, удлинение хромосом и выявление аксиальных гранул связаны с дифференциальной деконденсацией хромосом, в результате чего менее устойнчивый к гипотоническому воздействию материал R-бэндов деконденсируется, тогда как матерал G-бэндов остается относительно более компактным. Мы предполагаем, что данный эффект может быть связан с частичной экстракцией
16
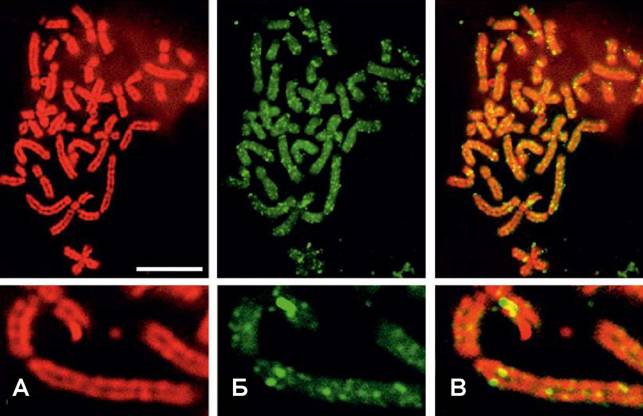
Рис. 6. Репликативные сайты, соответствующие позднореплицирующимся доменам хроматина, демонстрируют преимущественно осевую локализацию в хромосомах, полученных методом двойной гипотонической обработки. (А) ДНК окрашена бромистым этидием (красный), (Б) места включения БрдУ (зеленый), (В) наложение. Масштаб: 10 мкм.
и/или перераспределением структурных белков хроматина. Необычный ответ митотических хромосом, образовавшихся в процессе компактизации такого структурно модифицированного хроматина, на повторное гипотоническое возндействие показывает, что эти изменения хроматина сохраняются в течение иннтерфазы и сказываются на характере компактизации или структурной стабильнности хромосом в митозе.
Окрашивание таких хромосом бромистым этидием выявляло отчетливо видимое гало ДНК-содержащего материала, окружающее сестринские хромати-ды. Такое гало выявлялось исключительно в препаратах полученных после двойной гипотонической обработки. При анализе топологии рано- и поздно реплицирующегося хроматина в дифференциально деконденсированных хронмосомах обнаружилось, что большинство рано реплицирующихся сайтов вынявлялось в периферическом гало, окружающем хромосомы, или в пространстве между двумя сестринскими хроматидами, тогда как поздно реплицирующиеся домены располагались в осевых районах хромосом. Двойная гипотоническая обработка, вызывающая частичную деконденсацию хромосом, приводит также
17
к пространственному разобщению индивидуальных репликативных сайтов, вынявляя гетерогенную структуру таких доменов. Таким образом, полученные нами данные вполне соответствуют гипотезе о композитной организации мнонгих больших репликативных сайтов (Berezney et al. 2000, Koberna et al, 2005). В контрольных препаратах, приготовленных по стандартной методике, рано- и поздно реплицирующиеся домены демонстрировали одинаковое расположение относительно хромосомной оси.
Причина поперечной (и продольной) дифференцированности хромосом может состоять в различиях молекулярной организации G- и R-бэндов, в частности, различные по составу ансамбли структурных белков хроматина монгут обеспечивать компактизацию ДНК рано- и позднореплицирующихся районнов хромосом, функционируя в качестве скрепок, стабилизирующих высшие уровни организации хромосом. Выход рано реплицирующеся хроматина в гало при двойной гипотонической обработке может быть следствием более высокой чувствительности скрепок, поддерживающих структуру R-бэндов, к гипотонническому воздействию. В качестве возможных кандидатов на роль таких скренпок можно рассматривать конденсины. Поскольку распределение конденсинов I и II отчасти симулирует продольную дифференцированность хромосом (Опо et al, 2004), можно предположить, что они (или другие структурные белки, прянмо или опосредованно рекрутируемые конденсинами (Hudson et al, 2003)), определяют поперечную дифференцированность хромосом и различия в пронстранственной организации хроматина G- и R-бэндов в деконденсированных хромосомах.
3.4.а Динамикаа высшиха уровнейаа организацииа хроматинаа при активации транскрипции.
Для исследования поведения хроматиновых доменов высшего порядка при активации транскрипции была разработана модельная система, позволяюнщая визуализовать искусственные хромосомные локусы, содержащие индуци-бельные гены, при помощи взаимодействия 1ас-оператор/1ас-репрессор. Для этого были созданы генетические конструкции, в которых 256 копий тандемных повторов лак-оператора и ген устойчивости к канамицину/неомицину были фланкированы 19 п.н. сайтами узнавания транспозазы Тп5. Транспозицию даннных конструкций в ВАС-клоны, содержащие гены дигидрофолат-редуктаза (DHFR) мыши и человеческие гены металотионеина и белка теплового шока Hsp70, существляли in vitro при помощи очищенной транспозазы Тп5 и транснформировали рекомбинантные хромосомы в E.coli. Сайты интеграции транспо-зонов определяли путем секвенирования ДНК.
18
Таблица 1. Изменение уровня экспрессии трансгенных конструкций при
активации транскрипции
HSP705 |
||||
контроль |
Тепловой шок |
|||
эндогенный |
трансген |
эндогенный |
трансген |
|
Относительный уровень мРНК |
1 |
0.60.11 |
14023 |
20056 |
На одну копию |
1 |
0.140.02 |
14023 |
4512 |
МТ1_1_4 |
DHFRD102 |
||||
контроль |
Zn+2 2 часа |
Zn+2а 16 ч |
Контроль (log-фаза) |
GO |
|
Относительный уровень мРНК |
1 |
2.90.14 |
518.3 |
1 |
0.310.046 |
Каждую рекомбинантную ВАС трансфицировали в клетки СНО DG44, стабильно экспрессирующие химерный белок EGFP- димер лак-репрессора-NLS, отбирали стабильные клоны, содержащие несколько интегрированных конпий трансгена на ядро, располагающихся в виде серии GFP-позитивных точек.
Преимущества разработанного нами экспериментального подхода с иснпользованием ВАС состоят в том, что он дает возможность изучения структурнных преобразований высших уровней организации хроматина при активации транскрипции в системе, наиболее точно соответствующей поведению натив-ных генов в их естественном хроматиновом контексте. Сравнение уровней экснпрессии и степени индукции транскрипции эндогенных генов и ВАС трансгеннов, при помощи количественной РТ-ПЦР с использованием видо специфичных праймеров, при перерасчете на одну копию трансгена, показало, что после инндукции эндогенные и трансгенные копии генов демонстрируют сходные уровни активности (см. таблицу 1).
Параллельно мы провели сравнительный анализ структурной организации высших уровней компактизации хроматина до и после индукции транскрипции во всех трех трансгенных линиях. Для этого на светооптическом уровне были измерены изменение числа GFP-позитивных точек, а также определена контурная длина кривой, соединяющей все точки (см. таблицу 2). Оказалось, что средняя степень компактизации в неиндуцированных трансгенных локусах была в 25-50 раз выше, чем расчетный линейный коэффициент компактизации 30-нм фибриллы хроматина. 1.5-3-кратная деконденсация, наблюдаемая в результате индукции транскрипции, дает в результате коэффициент компактизации около 175, что все еще в несколько раз выше, чем компактизация 30-нм фибриллы. Полученные данные не исключают возможности того, что активно транскрибирующеся гены в составе трансгенной
19
Таблица 2. Изменения степени компактизации трансгенных локусов при активации транскрипции
Трансгенная линия |
DHFRD102 |
Hsp70_5 |
МТ114 |
Копийность (QPCR) |
13(10-16) |
10(8-14) |
9(7-11) |
Копийность (Fiber-FISH) |
11/12/11 |
9-12 |
16-18 |
Размер ВАС (т.п.н) |
187.1 |
192.6 |
185.2 |
Средняя контурная длина локуса (контроль, мкм) |
0.65 |
0.52 |
0.69 |
Коэффициент компактизации (контроль) |
1125 |
1200 |
1550 |
Средняя контурная длина локуса (после активации, мкм) |
1.4 |
1.51 |
0.98 |
Коэффициент компактизации (после активации) |
520 |
410 |
1090 |
конструкции образуют сильно деконденсированные петли ДНК, выпетливающиеся наружу из высоко конденсированной коровой фибриллы хроматина высшего порядка (хромонемы), содержащей повторы лак-оператора. Для проверки этой возможности был проведен анализ взаимного расположения повторов лак-оператора и ВАС методом двухцветной FISH. Метка лак-оператора обычно локализовалась колинеарно с последовательностями ВАС и обычно демонстрировала высокую степень колокализации с последней. Мы не наблюдали выпетливания участков ДНК ВАС на заметное расстояние от линейного лостова трансгенного локуса (рис.7).
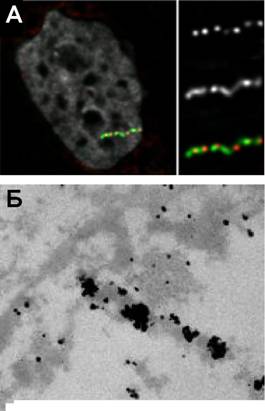
Далее мы использовали метод in vivo иммуномечения для визуализации структурной организации DHFR трансгенов в клетках линии В10 и В9, используя моноклональные антитела против GFP В полном соответствии с наблюдениями, полученными при помощи 3D-FISH, мы обнаружили локусы лак-репрессора в составе структур высшего конденсированного хроматина, соответствующих элементам хромонемы толщиной 100-130 нм (рис. 7).
Чтобы выяснить, является ли транскрипционая активность гена необходимым условием декомпактизации высших уровней упаковки хроматина, был проведен ингибиторный анализ поведения трансгенов, содержащих Hsp70 или МТ ВАС с использованием альфа-аманитина и 5,6-
20
-200 nm
Рис.7. А). Повторы лак-оператора и участки ВАК, сондержащие транскрибирующиенся гены, сохраняют колинеарнность при активации транснкрипции., Б) Хроматин, содернжащий транскрибирующиеся локусы, сохраняет хромонемную структуру.
дихлоро-1-бета-0-рибобензимидазола
(DRB). При действии альфа-аманитина
даже в условиях активации транскрипции
трансгенных локусов степень их
компактизации несколько увеличивается
по сравнению с неиндуцированными
контрольными клетками, что можно
объяснитьа ингибированием
конститутивного уровня экспрессии. При
использованииаа DRB степень
деконденсации была существенно ниже, чем в необработанных DRB клетках при индукции Hsp70. Для МТ ВАС статистически достоверных отличий в степени компактизации трансгенного локуса в контрольных неиндуцированных клетках и в клетках, обработанных Zn в присутствие DRB, обнаружено не было. Поскольку ген Hsp70 находится в активированном состоянии и с ним постоянно связана РНК полимераза II (Price, 2008), различия в поведении трансгенов при действии этих ингибиторов могут быть связаны с наличием РНК полимеразы на промоторе гена Hsp70. Присутствие DRB, не влияющее на связь РНК полимеразы с промотором, таким образом, не препятствует инициации транскрипции после теплового шока.
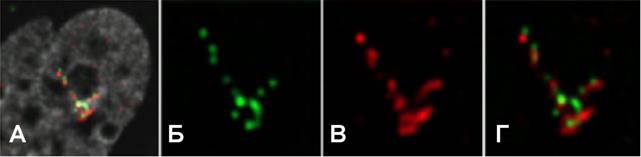
Рис.8. РНК-FISHс зондами против гена DHFR(В) показывает, что практически все копии в составе тандемных повторов DHFRВАС, интегрировавшихся в хромосому, являются транскрипционно-активными.(Б) Lac-penpeccop-GFP, (Г) совмещение.
21
Поскольку уровень экспрессии исследованных трансгенов оказывается в несколько раз ниже, чем у соответствующих эндогенных генов, мы оценили долю активных генов в составе исследуемых трансгенных локусов при помощи РНК-FISH. Гибридизационный сигнал при этом выявлялся в тесной ассоциации с большинством GFP-позитивных сайтов локализации повторов лак-оператора. Также часто наблюдалась частичная или полная колокализация гибридизационного и GFP-сигналов. Таким образом, активация транскрипции, по-видимому, происходит в большинстве индивидуальных копий трансгена вне зависимости от локальной степени компактизации. Также PHK-FISH-анализ показал, что экспрессия Hsp70 начинается в 90% клеток через 5 мин после начала теплового шока, тогда как деконденсация высших уровней компактизации хроматина трансгенного локуса наблюдается лишь спустя 10-30
|
мин.
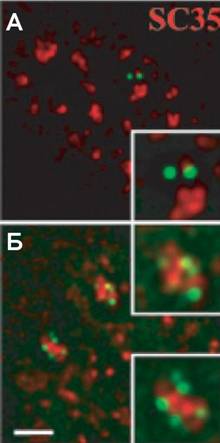
Рис. 9. Активация траннскрипции трансгенного локуса, содержащего гены теплового шока, стимулинрует ассоциацию транснгена (зеленый) с класнтерами интерхроматинонвых гранул (красный). До (А) и после активации (Б) |
При индукции трансгенные локусы DHFR и МТ-ВАС как правило принимали линейную конфор-мацию, гда как в случае активации гена Hsp70 тепловым шоком в течение 30 мин значительная часть Hsp70 трансгена приобретала специфиченскую форму полукруга. Более того, при длительнных тепловых воздействиях (30-40 мин) транснкрипты аккумулировались в ДАПИ-негативной области, которую охватывала цепочка GFP-позинтивных точек (рис. 6). Иммунофлуоресцентная микроскопия показала, что Hsp70 трансгены, в отнличие от DHFR и МТ трансгенов, тесно связаны с ядерными доменами, соответствующими кланстерам интерхроматиновых гранул (nuclear speckles), окрашиваемыми антителами против SC-35 и ASF/SF2 (рис.9). Такая ассоциация характернна для всех независимо полученных клеточных линий, содержащих Hsp70 трансген (но не для DHFR и МТ-трансгенов). In vivo иммуномечение золотом лак-репрессора-GFP в составе Hsp70 ВАС трансгена демонстрирует расположение повторов лак-оператора на периферии кластеров интерхронматиновых гранул. Анализ пространственного расположения трансгенов относительно кластера интерхроматиновых гранул на стереоизображенинях срезов толщиной 0.5 мкм или на сериях нанклонных проекций показывает их локализацию в области аморфного фибриллярного материала, окружающего плотный центральный гранулярный домен. Сочетание иммунофлуоресценции и РНК-
22
FISH подтверждает, что аккумуляция транскриптов Hsp70 происходит внутри SC-35-позитивных ядерных доменов на более поздних сроках после начала тепнловой обработки. Индекс ассоциации увеличивался с 47% до 85% в течение 30 мин после теплового шока, тогда как средний размер ближайшего кластера оставался относительно стабильным. Это можно объяснить перемещением Hsp70 трансгена к уже существующему кластеру. Чтобы исследовать это явленние более подробно, была проведена серия экспериментов по прижизненному наблюдению за клетками в процессе активации гена теплового шока. Для этой цели клетки, несущие трансгенную конструкцию Hsp-70 ВАС и экспрессирую-ще GFP-flHMep-Lac-penpeccopa, были трансформированы плазмидой, кодируюнщей mRFP-ASF/SF2.
|
Страницы: | 1 | 2 | 3 | 4 | 5 | 6 | |
Авторефераты по всем темам >>
Авторефераты по биологии