
Н. Е. Булыгин дендрология 2-е издание. Переработанное и дополненное допущено Государственным комитетом СССР по на- родному образованию в качестве учебник
| Вид материала | Учебник |
СодержаниеСистематика и характеристика покрытосеменных 5.2. Древесные растения подкласса магнолииды 5.3. Древесные растения подкласса ранункулиды Барбарис амурский Барбарис сибирский |
- В. М. Смолевского Издание 3-е, переработанное и дополненное Допущено Государственным, 5319.11kb.
- А. И. Соловьев введение в политологию Издание третье, переработанное и дополненное, 6711.57kb.
- А. И. Соловьев введение в политологию Издание третье, переработанное и дополненное, 6532.51kb.
- А. Г. Кучерена адвокатура второе издание, переработанное и дополненное Допущено Учебно-методическим, 12778.36kb.
- Учебник 3-е издание, переработанное и дополненное, 10138.23kb.
- А. Г. Асмолов психология личности принципы общепсихологического анализа Допущено Государственным, 4733.92kb.
- Учебник издание пятое, переработанное и дополненное проспект москва 2001 Том 3 удк, 11230.01kb.
- Учебник издание пятое, переработанное и дополненное проспект москва 2001 Том 3 удк, 11433.24kb.
- Учебник. 3-е издание, переработанное и дополненное, 10586.44kb.
- Наших дней, 5300.94kb.
СИСТЕМАТИКА И ХАРАКТЕРИСТИКА ПОКРЫТОСЕМЕННЫХ
5. 1. Общая характеристика отдела покрытосеменные
(Magnoliophyta, или Angiospermae)
Покрытосеменные, или цветковые, растения представляют со-бой самый обширный отдел растительного мира. Он включает не менее 240 000 видов, относящихся примерно к 13 000 родам свыше 390 семейств. По числу видов покрытосеменные значительно пре-восходят все остальные группы высших растений, взятые вместе.
Как особая ветвь эволюции, покрытосеменные произошли от семенных папоротников около 120 млн лет назад, в нижнемело-вую эпоху. Большинство современных систематиков-эволюциони-стов (акад. Тахтаджян А. Л., 1970, 1987 и др.) считают, что наи-более вероятной прародиной, первичным центром расселения цвет-ковых были горные районы субтропиков Юго-Восточной Азии. В раннем мелу покрытосеменные еще не играли какой-то замет-ной роли в растительном покрове Земли. Однако в середине ме-лового периода (примерно 100 млн лет назад) происходит одно из наиболее глубоких и резких изменений растительного мира суши, и цветковые за сравнительно короткий период геологиче-ского времени широко распространяются по земному шару, до-стигнув Арктики и Антарктики.
В современную геологическую эпоху цветковые растения про-израстают во всех климатических зонах и в самых разных эколо-гических условиях - от тропических лесов до тундр, от болот до пустынь и от морских побережий до высокогорий. Они составляют основную массу растительного вещества биосферы и являются самой важной для человека группой растений.
Одним из основных условий столь бурного распространения цветковых явилась их высокая эволюционная пластичность, про-явившаяся в необычайном разнообразии многочисленных при-способлений к самым различным условиям внешней среды. От голосеменных цветковые отличаются прежде всего тем, что семязачатки (семяпочки) у них заключены в более или менее замкнутую полость завязи, образованной одним или несколькими сросшимися плодолистиками. Поэтому цветковые растения чаще
называют покрытосеменными. Благодаря тому, что семязачатки заключены в полость, у покрытосеменных пыльца попадает не непосредственно на микропиле семязачатка, а на рыльце.
Наличие рыльца является характернейшей особенностью цвет-ковых растений и фактически главным отличием цветка от стро-била голосеменных. Весьма характерны также гаметофиты цвет-ковых растений, которые до крайности редуцированы, что позво-ляет им развиваться более ускоренными темпами, чем гаметофиты голосеменных. Совершенным стал и процесс оплодотворения: вза-мен простого, присущего голосеменным, у покрытосеменных про-исходит оплодотворение двойное, впервые открытое академиком С. Г. Навашиным.
Суть этого сложного биологического процесса состоит в том, что один из двух образующихся в мужском гаметофите спермиев (мужских гамет) сливается с яйцеклеткой (собственно оплодо-творение), а другой - с двумя свободными или уже слившимися так называемыми полярными ядрами женского гаметофита (трой-ное слияние). В результате слияния одного из спермиев с яйце-клеткой образуется зигота; в результате же тройного слияния -первичное ядро эндосперма, который представляет собой харак-терную для цветковых растений питательную ткань, служащую для питания развивающегося из зиготы зародыша.
Биологическое значение двойного оплодотворения состоит в том, что эндосперм (вторичный) у покрытосеменных развивается только при условии оплодотворения яйцеклетки. Если же оплодо-творение по каким-либо причинам не происходит, то и эндосперм не развивается. Таким образом исключается напрасная трата пи-тательных веществ на образование эндосперма в случае отсут-ствия его потребителя - зародыша семени, не развивающегося из-за нарушенного оплодотворения. Простое оплодотворение го-лосеменных в этом отношении менее совершенно, так как при про-стом оплодотворении эндосперм образуется до оплодотворения независимо от того, формируется ли зародыш или нет.
В результате двойного оплодотворения эндосперм покрытосе-менных несет в себе наследственные задатки как материнского, так и отцовского организмов. Весьма вероятно, что именно в этом кроется одна из причин той необычайно высокой приспособлен-ности покрытосеменных к самым различным экологическим усло-виям, которую они проявили в процессе эволюции. Голосеменные, эндосперм которых имеет задатки только материнской наслед-ственности, таким широким диапазоном приспособительных воз-можностей не обладает.
В процессе эволюции у цветковых появился и такой новый ор-ган, как околоцветник. Он выполняет целый ряд важных вспомо-гательных функций: защиту пестика и тычинок от неблагоприят-ных воздействий внешней среды, привлечение насекомых или птиц - посредников опыления и др. В частности, быстрому рас-
пространению и возникновению множества биологических форм покрытосеменных способствовало появление насекомых-опылите-лей, период массового развития которых совпал с интенсивным заселением суши растениями этого отдела. Немаловажное зна-чение тут имели и птицы - активные разносчики пыльцы и семян.
В отличие от голосеменных для покрытосеменных растений характерно наличие в ксилеме наряду с трахеидами также сосу-дов. Значительные различия имеются и в строении флоэмы: си-товидные элементы всех цветковых растений снабжены так назы-ваемыми клетками-спутницами, отсутствующими у голосеменных.
Чрезвычайно высокую способность к изменчивости проявили листья покрытосеменных. Они настолько сильно отличаются от игловидных и чешуевидных листьев хвойных, что это послужило основанием подразделять древесные растения на хвойные и лист-венные породы.
По сравнению с голосеменными у покрытосеменных появился целый ряд новых биоформ (кустарнички и полукустарнички, по-лукустарники, травы), более широкое развитие получили вегета-тивное размножение и возобновление, а успешной адаптации к сезонным изменениям климатических условий различных природ-ных зон способствовала выработка многообразных феноритмо-типов и фенологических форм, не свойственных голосеменным (см. ниже характеристику лиственных древесных растений). В це-лом же цветковые растения достигли более высокого уровня эво-люционного развития, чем голосеменные.
Все покрытосеменные подразделяются на два класса: двудоль-ные и однодольные.
Класс двудольные (Magnoliopsida) характеризуется наличием двух семядолей у зародыша семени, открытыми проводящими пучками, сохранением в течение всей жизни главного корня, сет-чатым жилкованием листьев, пяти-, четырех-, двух- или много-членным типом строения цветка. К этому классу относятся не ме-нее 180 000 видов покрытосеменных растений.
Класс однодольные (Liliopsida) отличается наличием одной семядоли у зародыша, закрытыми (без камбия) проводящими пучками, ранним отмиранием главного корня и развитием прида-точных корней, параллельным или дугонервным жилкованием листьев, трехчленным строением цветка. Имеющиеся среди однодольных древовидных формы вторичного происхождения характеризуются совершенно иным, чем у двудольных, способом утолщения ствола. Однодольные включают около 60 000 видов. Большинство систематиков считают, что они произошли от дву-дольных еще на ранней ступени эволюции, а в дальнейшем оба класса развивались независимо и параллельно.
Из многочисленных филогенетических (естественных) систем покрытосеменных растений, созданных в XVIII-XX вв. как отечественными (Горяников П. Ф., Кузнецов Н. И., Буш Н. А.,
Козо-Полянский Б. М., Гроссгейм А. А., Тахтаджян А. Л.), так и зарубежными (Энглер А., Халлир X., Кронквист А., Торн Р., Дальгрен Р.) ботаниками, наиболее передовой является система, разработанная советским ботаником академиком А. Л. Тахтад-жяном (1966, 1970, 1987).
Согласно А. Л. Тахтаджяну, класс двудольные включает 7 под-классов: Magnoliidae, Ranunculidae, Hamamelididae, Caryophyl-lidae, Dilleniidae, Rosidae, Asteridae. Однодольные подразделя-ются на три подкласса: Alismatidae, Liliidae и Arecidae.
В пределах каждого подкласса его семейства объединены в порядки, порядки - в надпорядки. Двудольные включают 71 по-рядок, 20 надпорядков, однодольные - 21 порядок, 8 надпоряд-ков. Первые по счету надпорядки охватывают наиболее прими-тивные порядки, последние - филогенетически более продви-нутые.
С филогенетической системой А. Л. Тахтаджяна, с графиче-ским изображением филогенетического древа покрытосеменных студенты уже знакомились в курсе ботаники. Поэтому отметим только, что среди покрытосеменных древесных и полудревесных растений насчитывается около 100 000 видов. Из них свыше 92 000 относятся к двудольным. У однодольных древовидные фор-мы (вторичного происхождения) наиболее полно представлены в порядках пальмы (Arecales) - примерно 3000 видов, пандано-вые (Pandanales) - свыше 800 видов и некоторых других.
В учебнике охарактеризованы покрытосеменные древесные растения 303 видов и 25 форм (культиваров) класса двудольные. Они относятся к 112 родам 41 семейства 7 подклассов. Подклассы и порядки изучаемых семейств указаны в табл. 2.
Следует заметить, что во многих отечественных и зарубежных справочниках и дендрологических сводках при характеристике покрытосеменных древесных растений использована филогене-тическая система А. Энглера (Деревья и кустарники СССР. 1951 -1962, Т. 2-6; География древесных растений СССР. 1965; Ареалы деревьев и кустарников СССР. 1977-1986. Т. 1-3 и др.). В ряде учебников по дендрологии (Гроздов Б. В., 1952, 1960; Бог-данов П. Л., 1974) нашла отражение система покрытосеменных растений, предложенная А. А. Гроссгеймом. Обе эти устаревшие системы существенно отличаются от принятой в данном учебнике современной филогенетической системы академика А. Л. Тах-таджяна.
Для более полной характеристики цветков в тексте часто упо-требляется их ботаническая формула. Поэтому напомним, что морфологически различные круги цветка в формуле принято обо-значать начальными буквами их латинских или греческих назва-ний: Р - простой околоцветник, К - чашечка, С - венчик, А -тычинки, G - пестик(и). Число членов цветка указывают цифрой
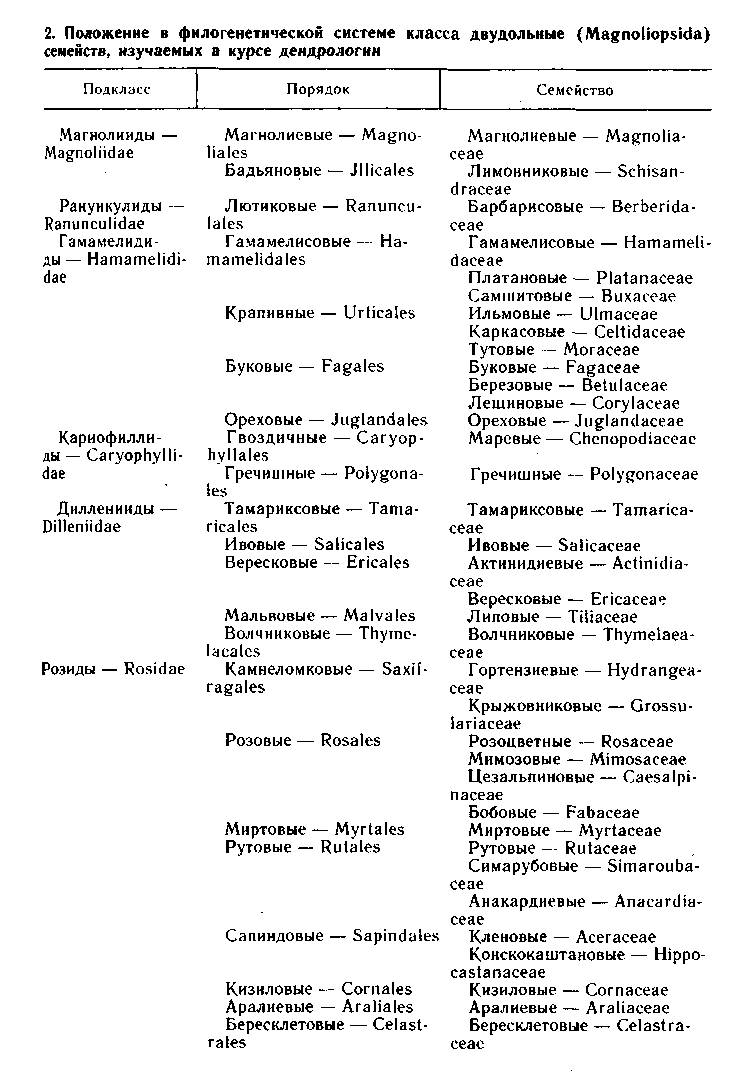
или знаком бесконечности, если их более 10, а при отсутствии чле-нов в данном круге - нулем.
Сращение членов показывают заключением цифры (знака бесконечности) в круглые скобки; расположение частей цветка в несколько кругов - знаком плюс; положение завязи в цветке обо-значают чертой: при верхней завязи черту проводят под цифрой числа плодолистиков; при нижней завязи - над ней, при полу-нижней - посередине, справа от цифры; цветки актиноморф-ные *; зигоморфные ??? ; тычиночные ♂ ;пестичные ♀ .
5.2. Древесные растения подкласса магнолииды
(Magnoliidae)
Среди растений этого подкласса доминируют древесные -свыше 70 % таксономического состава, или около 7100 видов. У целого ряда видов сосуды еще отсутствуют. Как правило, цвет-ки обоеполые, спиральные или спироциклические. Гинецей в боль-шинстве случаев апокарпный (т. е. состоит из свободных плодо-листиков) . Семена обычно с мелким зародышем, с обильным эндо-спермом, а иногда и с периспермом.
Представители семейства магнолиевые (Magnoliaceae) распространены преимущественно в субтропических областях Се-верного полушария. Вечнозеленые, реже листопадные (зимнего-лые) деревья и кустарники с простыми, очередными, иногда очень крупными (до 1 м длиной) листьями. Цветки одиночные, верхушеч-ные или пазушные, обычно крупные, с простым многочисленным околоцветником, кремово-белые, розовые, зеленовато-желтые, обоеполые или раздельнополые, с многочисленными тычинками и пестиками, расположенными спирально на удлиненном цвето-ложе. Опыляются определенными видами жуков. Плод - сбор-ная листовка, внешне напоминающая шишку хвойных (рис. 18).
Род магнолия (Magnolia) насчитывает около 70 видов, из которых в СССР естественно растет только один - магнолия об-ратнояйцевидная, или белоспинная (М. obovata). Листопадное дерево 8-12 м высотой; побеги с восковым налетом, листья об-ратнояйцевые (рис. 18), 20-40 см длиной и до 20 см шириной,

Рис. 18. Магнолия белоспинная:
1 - лист; 2 - цветок; 3 - плод; 4 - семя
цельнокрайние; цветки кре-мово-белые, до 18 см в диаметре(* P∞A∞G∞), цветут вскоре после рас-пускания листьев; плод-эллиптическая, ярко-крас-ная сборная листовка, 12-18 см длиной. Растет маг-нолия белоспинная только на Южно-Курильских ост-ровах и как редкий вид занесена в «Красную кни-гу СССР». Весьма декора-тивна, заслуживает широ-кого использования в озе-ленении, особенно в При-морье Дальнего Востока.
На Черноморском побережье Кавказа и Крыма обычна в пар-ковой культуре магнолия крупноцветковая (М. grandiflora), интродуцированная из Северной Америки. Крупное (до 30 м вы-сотой и свыше 1 м в диаметре ствола), вечнозеленое дерево с об-ратнояйцевидными или узкоэллиптическими листьями 12-25 см длиной. Листья цельнокрайние, кожистые, с верхней стороны темнозеленые, глянцевые, с нижней - рыжеватые от короткого опушения. Цветки молочно-белые, до 22 см в диаметре, с сильным одуряющим запахом, распускаются в кроне постепенно, отчего цветение продолжается долгое время. Плоды шишковидные, до 12 см длиной и 4 см толщиной; семена черные, блестящие, оваль-ные, 6-8 мм длиной, при созревании повисают на красных ни-тях - семяносцах. Из-за низкой зимостойкости м. крупноцветко-вая перспективна для озеленения только в районах советских субтропиков.
Род лириодендрон, или тюльпановое дерево (Li-riodendron). Представителем этого рода является лириодендрон тюльпанный (L. tulipifera) - крупное дерево североамерикан-ских лесов с ценной древесиной. Листья четырехлопастные, с выемкой на вершине, 12-15 см длиной. Цветет в первой полови-не - середине лета, цветки чашеобразные, около 6 см в диаметре (* Р3+3+3 А∞G∞), зеленовато-оранжевые. Плод сборный, уз-коцилиндрический, состоит из многочисленных двухсемянных кры-латок, до 3,5 см длиной, сидящих на разросшемся цветоложе. Де-

Рис. 19. Лимонник китайский: ветви с цветками и с соплодием
рево ценное для озеленения. В СССР используется преиму-щественно в Закавказье.
Виды семейства лимон-никовые (Schisandraceae) обитают преимущественно в Во-сточной и Юго-Восточной Азии. Из них в хвойно-широколист-венных лесах Дальнего Восто-ка естественно распространен только один вид рода лимон-ник (Schisandra) -лимонник китайский (Sch. chinensis). Это вьющаяся листопадная лиана 4-8 (15) м длиной. Листья оче-редные, эллиптические или об-ратнояйцевидные,5-10 см дли-ной, до 5 см шириной, с кли-
новидным основанием, постепенно заостренные к вершине, по краю неясно-зазубренные, сверху темно-зеленые, снизу блед-нее, по жилкам слабо опушены; черешок красноватый, до 3 см длиной. На укороченных плодоносных веточках листья собра-ны по 2-3 в пучке. Цветки воронковидные, до 2 см в диаметре, белые, раздельнополые (6'*КзСбАсо; $*К.зСбОм), душистые, свисают на длинных розоватых цветоножках. Растение многодом-ное, цветет после облиствения. Плоды сборные, состоят из много-численных красных двухсемянных ягод, расположенных на удли-ненном цветоложе, напоминая кисть (рис. 19), созревают в на-чале осени. Семена почковидные, желто-оранжевые, около 3 мм в диаметре. Стебли, корни и особенно плоды при растирании издают своебразный аромат, напоминающий запах лимона (отсюда и на-звание растения). Плоды кисловатые, сильновяжущие, обладают высокотонизирующим действием, поэтому широко используются в медицине. Но лимонник ценен и для вертикального озеленения.
Размножают его семенами, отводками, корневыми отпрысками, зелеными (летними) черенками. Лимонник сравнительно теневы-нослив, среднетребователен к плодородию и влажности почвы, довольно зимостоек. Успешно культивируется в европейской ча-сти СССР, включая Карельский перешеек, где может обильно плодоносить.
Семейство лавровые (Lauraceae) - это преимущественно растения тропиков и субтропиков. Из представителей этого се-мейства в одичалом виде получил распространение в Закавказье
один из видов рода лавр (Laurus) - лавр благородный (L. nо-bilis) - интродуцированный из Средиземноморья. Вечнозеленое теплолюбивое дерево до 18 м высоты, нередко кустовидное. Листья очередные, простые, цельнокрайние, но слегка волнистые, про-долговатые или ланцетные, 8-20 см длиной, 2-8 см шириной, плотнокожистые. Цветки мелкие, в пазушных зонтиках, раздель-нополые или обоеполые. Плод черный, ягодовидный, при осно-вании окружен разрастающимся цветоложем. Лавр широко разводят на Кавказе и Южном берегу Крыма как ценное эфиро-масличное, пряное и высокодекоративное растение. Его листья являются объектом промышленной заготовки и используются в качестве приправы. Лавр хорошо переносит стрижку и пригоден для создания живых изгородей.
5.3. Древесные растения подкласса ранункулиды
[Ranunculidae]
Среди этого подкласса древесных и полудревесных растений насчитывается около 1700 видов. Все представители подкласса обладают сосудами. Цветки обоеполые или однополые, часто спи-ральные или спироциклические. Семена обычно с мелким заро-дышем и хорошо развитым эндоспермом, хотя могут быть и без эндосперма. Предполагают, что ранункулиды произошли непо-средственно от магнолиид, скорее всего, от предков типа бадь-яновых.
Виды семейства барбарисовые (Berberidaceae) распро-странены преимущественно в умеренных и субтропических обла-стях Северного полушария. Сравнительно-анатомические иссле-дования показывают, что древесные барбарисовые произошли от травянистых предков.
В естественной дендрофлоре СССР барбарисовые представ-лены родом барбарис (Berberis), который насчитывает около 500 видов, в СССР дико растут 18, интродуцированы свыше 60.
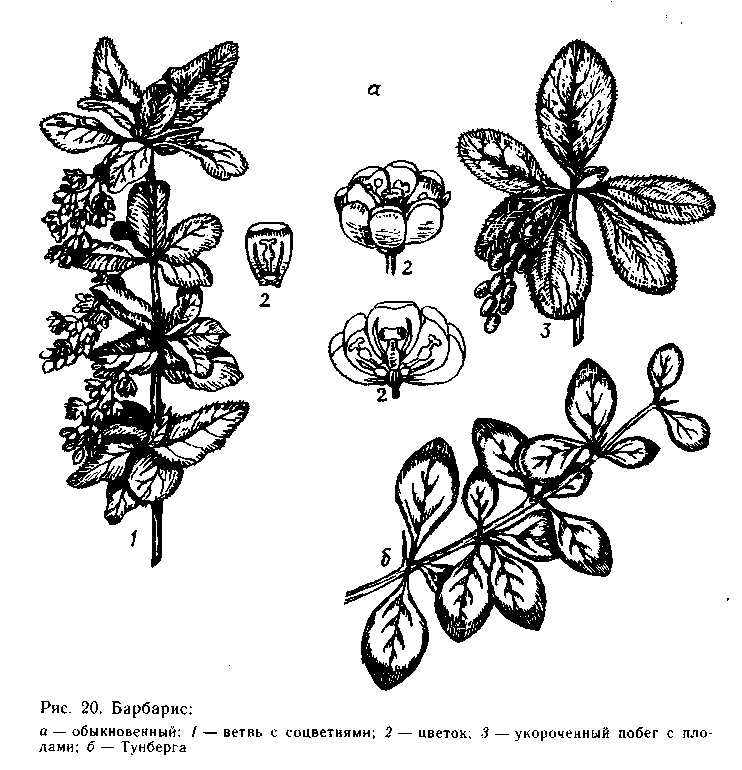
Барбарисы - вечнозеленые, полувечнозеленые или листопад-ные кустарники, реже невысокие деревья с тонкими, обычно реб-ристыми удлиненными побегами и образующимися на них много-численными брахибластами. На удлиненных побегах листья оче-редные, на укороченных - собраны в пучки. Листья простые, от ланцетных до обратнояйцевидных, голые, цельнокрайние, мелко-реснитчатые или колючезубчатые, на коротком черешке. Прилист-ники неопадающие, листоватые, слабозубчатые или превращен-ные в колючки. Цветки в поникающих кистях, пучках или одиноч-ные на концах брахибластов, мелкие, желтые, с нектарниками (*К6С6А6G1), цветут после распускания листьев. Плод - эл-липтически-шаровидная ягода, 8-12 мм длиной, черная или красная, с остающимся засохшим рыльцем, с 1-5 продолгова-тыми, коричневыми семенами. Виды барбариса образуют плот-
ную густую крону, хорошо переносят стрижку, ценятся в озеле-нении. Их прочная древесина идет на мелкие поделки, из коры и корней получают желтую краску для окраски кож. Листья и пло-ды барбариса кислые от большого содержания яблочной кислоты, используются в пищевой промышленности и населением для при-готовления приправ и настоек.
В СССР наиболее широко распространен барбарис обыкно-венный (В. vulgaris). Естественно он растет в хвойно-широко-лиственных и широколиственных лесах европейской части, в Кры-му и на Кавказе. Листопадный, раскидистый кустарник до 2,5 м высоты. Листья обратнояйцевидные, до 4 см длиной и 2 см ши-риной, к основанию сужены, с закругленной или заостренной вер-хушкой, по краю мелко- и острозубчато-пильчатые (рис. 20). Ко-
лючки обычно 3-5-раздельные, до 2 см длиной. Цветки в кистях желтые, 6-9 мм в диаметре. Ягоды продолговато-эллиптические, до 12 мм длиной, пурпурные, семена 4-5 мм длиной, слегка сплюс-нутые, темно-коричневые.
Барбарис среднетребователен к плодородию и влажности поч-вы, теневынослив, довольно зимостоек, обычно отличается ран-ним началом и поздним завершением вегетации.
Очень эффектен пурпурный культивар барбариса обыкновен-ного - В. vulgaris 'Atropurpurea'. Чашелистики его цветков темно-красно-фиолетовые, листья весной темно-пурпурные, поз-же - пурпурно-сизоватые. Оригинальная окраска этого культи-вара только частично передается потомству при семенном раз-множении.
Барбарис амурский (В. amurensis) - вид, замещающий б. обыкновенный в лесах Приморья Дальнего Востока. Отличается большей высотой - до 3,5 м, довольно жесткими листьями до 12 см длиной, мощными, 3-5 (7)-раздельными колючками до 3 см длиной.
Барбарис сибирский (В. sibirica) - листопадный кустарник до
- м высотой. Листья кожистые, продолговато-яйцевидные до
- см длиной, цветки желтые, 10-12 мм в диаметре, одиночные, плоды красные. Распространён в горах Алтая, Саян и Средней Азии.
Из интродуцированных видов барбариса наибольшей попу-лярностью в озеленении в СССР пользуются барбарис Тунберга (В. thunbergii) родом из Японии. Кустарник до 1 м высотой, ред-ко до 2,5 м, побеги ярко-красные или красно-оранжевые, ветви ребристые, дугообразно отклоненные. Листья ромбически-оваль-ные, округлые или лопатчатые, с клиновидным основанием (вме-сте с черешком до 2 см длиной), по краю цельные (рис. 20), осенью, перед опадением, становятся пурпурово-малиново-фио-летовыми. Колючки мелкие, простые. Цветки в пучках или оди-ночные, желтые с красноватым оттенком, до 10 мм в диаметре. Плоды кораллово-красные, блестящие, эллипсоидальные, до 10 мм длиной. В экологическом отношении схож с барбарисом обык-новенным.
К этому же семейству относятся род м а г о н и я (Mahonia) -вечнозеленые кустарники с непарноперистосложными, очень же-сткими, темно-зелеными блестящими листьями. Цветки желтые, собраны в прямостоячие кисти, образующиеся в пазухах чешуй верхушечных цветковых почек. Цветут раньше барбарисов -до начала роста побегов. Ягоды продолговато-эллиптические, до 10 мм длиной, черные, с сизым налетом, созревают в конце лета -начале осени. Из видов этого рода в СССР наиболее часто встре-чается магония падуболистная (М. aquifolium) - кустарник до 1 м высотой, родом из Северной Америки.