
Естественные науки э. С. Велиляева влияние сленговых слов на энергоинформационную систему студентов горно-алтайского государственного университета
| Вид материала | Документы |
- Телекоммуникационная среда горно-алтайского университета и научно-образовательной сферы, 34.38kb.
- Учебное пособие «Политическая система кнр» для студентов специальностей «Регионоведение»,, 2761.81kb.
- «Горно-Алтайская станция скорой медицинской помощи», 19.89kb.
- Вестник Брянского государственного технического университета. 2007. №3(15) Естественные, 126.78kb.
- Название: Молодежь расспросили о программе, 27.41kb.
- Бюллетень новых поступлений литературы за 2011 год, 5843.31kb.
- Самостоятельная работа студентов методические рекомендации по выполнению самостоятельной, 922.35kb.
- Самостоятельная работа студентов методические рекомендации по дисциплине сд., 302.18kb.
- Рабочая программа по дисциплине «Физическая культура студентов специального медицинского, 2010.14kb.
- Владимиров Николаевичем Горбачевым. Они обсудили два блока проблем: экология Алтайского, 15.07kb.
О.И. Симонова, Т.Б. Баданова
ВЕНТИЛЯЦИОННО-ПЕРФУЗИОННЫЙ КОЭФФИЦИЕНТ У ДЕВУШЕК, ПРОЖИВАЮЩИХ В РЕСПУБЛИКЕ АЛТАЙ
Одной из актуальных задач физиологии и медицины является изучение роли соотношения вентиляции и перфузии газов в лёгких. Вопрос о механизмах развития изменений вентиляционно-перфузионного коэффициента при различных условиях представляет значительный теоретический и практический интерес.
Для нормального процесса обмена газов в легочных альвеолах необходимо, чтобы их вентиляция воздухом находилась в определенном соотношении с перфузией их капилляров кровью. Иными словами, минутному объему дыхания должен соответствовать минутный объем крови, протекающий через сосуды малого круга, а этот объем, равен объему крови, протекающий через большой круг. В обычных условиях вентиляционно-перфузионный коэффициент у человека составляет 0,8-0,9. Например, при альвеолярной вентиляции, равной 6 л/мин, минутный объем крови может составить около 7 л/мин.
В отдельных областях лёгких соотношение между вентиляцией и перфузией может быть неравномерным. Резкие изменения этих отношений могут вести к недостаточной артериализации крови, проходящей через капилляры альвеол. Легочные сосуды относят к «ёмкостной системе». Их просвет в значительной степени зависит от внутригрудного и внутриальвеолярного давления. В малом круге давление крови низкое, что в нормальных условиях предотвращает выпотевание плазмы через альвеолокапиллярную мембрану и образование отёка лёгких. Ширина сосудистого русла регулируется симпатической иннервацией. Имеются и местные механизмы, сопрягающие перфузию альвеол с их вентиляцией. Легочной кровоток в целом зависит от величины сердечного выброса, поэтому, в конечном счете, он управляется общими регуляторными механизмами сердечно-сосудистой системы. Отсюда тесная взаимосвязь между регуляцией дыхания и кровообращения, которая особенно ярко проявляется при мышечной деятельности. К этому следует добавить, что дыхательные колебания внутригрудного давления, действуя по принципу «двойного насоса», не только обеспечивают вентиляцию легких, но и помогают венозному возврату крови к сердцу. В свою очередь, пульсовые толчки давления в воздухоносных путях и альвеолах, вызванные сокращениями сердца, способствуют внутрилегочному смещению газов вызывая его кардиогенный компонент [3].
В различных климатогеографических зонах кардио-респираторная система организма проявляет приспособительные свойства, что вызывает особый интерес для её изучения, поэтому впервые с учётом зоны проживания была обследованы девушки Республики Алтай и проведена оценка значений вентиляционно-перфузионного коэффициента в покое и после дозированной нагрузки, как важного критерия функциональной активности кардиореспираторной системы.
Целью настоящей работы стало выяснение соответствия минутного объема крови минутному объему дыхания в покое и после дозированной нагрузки у девушек проживающих в разных климатогеографических зонах.
Для поставленной цели необходимо решить следующие задачи:
1.Рассчитать значения минутного объема крови и минутного объема дыхания до нагрузки у девушек, проживающих в разных климатогеографических зонах.
2.Рассчитать значения минутного объема крови и минутного объема дыхания после дозированной нагрузки.
3.Определить соответствие минутного объёма крови минутному объёму дыхания до и после дозированной нагрузки у девушек с учетом зоны проживания.
Объектом исследования являлись девушки, в количестве 30 человек проживающие в различных агроклиматических зонах. При процентном распределении девушек по зонам проживания использовалась классификация агроклиматических зон Республики Алтай предложенная Т.Д. Модиной, 1997 [2]. Большинство девушек проживали в низкогорье - 63%, в среднегорье проживало 23% и высокогорье - 13% обследованных.
У каждой девушки измеряли следующие физиометрические показатели в покое и после дозированной нагрузки: частоту сердечных сокращений (ЧСС), частоту дыхательных движений (ЧДД), систолическое артериальное давление (САД), диастолическое артериальное давление (ДАД). Подсчитывался минутный объем дыхания (МОД) и минутный объем крови (МОК) при этом использовались методики, приведённые в руководстве к лабораторным занятиям по общей и возрастной физиологии А.А. Гуминского [1].
По результатам исследования представленных в таблице 1, можно отметить, что в покое у девушек минутный объём крови был в среднем равным 5,5 литров, что соответствует стандартным значениям для данного возраста.
Минутный объём дыхания до нагрузки был равен у девушек, проживающих в высокогорье 4,9 литров, что достоверно ниже, чем у девушек, проживающих в среднегорье и ниже стандартных значений. После дозированной нагрузки минутный объём крови во всех группах увеличился на 1-1,5 л, а минутный объём дыхания увеличился в 5 раз. Коэффициент корреляции между показателями минутного объёма крови и минутного объёма дыхания был при этом положительным.
До нагрузки отмечалось высокая степень корреляции данных параметров, а после слабая. Результаты исследования представлены в таблице 1.
Таблица 1
| Показатели МОК и МОД до и после дозированной нагрузкиЗона проживания | МОК | r | МОД | r | ||
| До нагрузки (Х±m) (л/мин) | После нагрузки (Х±m) (л/мин) | До нагрузки (Х±m) (л) | После нагрузки (Х±m) (л) | |||
| высокогорье | 5,794±462,6 | 7,344±992,15 | 0,5 | 4,93±809 | 26,700±6,187 | 0,2 |
| среднегорье | 5,202±517,9 | 6,357±771,9 | 0,3 | 7,020±97 | 28,063±3,798 | 0,6 |
| низкогорье | 5,512±267,1 | 6,946±330,1 | 0,6 | 5,44±724 | 28,529±2,123 | 0,4 |
По результатам определения вентиляционно-перфузионного коэффициента можно отметить, что до нагрузки соответствие минутного объёма крови минутному объёму дыхания проявлялось у девушек проживающих в высокогорье и низкогорье. А после нагрузки это соответствие утрачивается. После дозированной нагрузки вентиляционно - перфузионный коэффициент резко увеличивается. Это объясняется тем, что кардиореспираторная система участников эксперимента не справляется, даже с незначительными физическими нагрузками, что по всей вероятности свидетельствует о малоподвижном образе жизни студенток, и развивающейся в следствие этого гиподинамии.
Выводы:
1. Значения минутного объёма крови и минутного объёма дыхания до нагрузки у девушек участвующих в исследовании соответствовали норме, кроме минутного объёма дыхания девушек проживающих в высокогорье (ниже нормы на 2 л), после нагрузки значения минутного объёма дыхания увеличивались на 1,5 л, а минутного объёма дыхания в 5 раз.
2. До нагрузки наблюдалось соответствие значений вентиляционно-перфузионного коэффициента, после нагрузки утрачивалось.
Литература
1. Гуминский, А.А. Руководство к лабораторным занятиям по общей и возрастной физиологии: Учеб.пособие для студентов биол. спец. пед. инстит-тов /А.А.Гуминский, Н.Н. Леонтьева, К.В. Маринова. – М.: Просвещение,1990. -239с.
2. Модина, Т.Д. Климат Республики Алтай/Т.Д. Модина. – Новосибирск, НПУ,1997. – 177с.
3. Ноздрачёв, А.Д. Начала физиологии: Учебник для вузов. /А.Д.Ноздрачёв, Ю.И. Баженов, И.А.Баранников, А.С. Батуев и др. /Под. ред. акад. А.Д. Ноздрачёва. – СПб.: Издательства «Лань», 2004 – 1088с.
Л.В. Шестакова
РЕЛИКТОВЫЕ ВИДЫ РОДА GALIUM L. ВО ФЛОРЕ РЕСПУБЛИКИ АЛТАЙ
При изучении любой флоры немаловажным моментом является анализ состава и возраста ее реликтов. Характерным признаком реликтовости видов является консервативность, не ограниченная изменчивость, часто несоответствие биологических особенностей вида современным условиям существования [1]. Изучению реликтов во флоре отдельных территорий посвящены многочисленные работы ученых [2–4] и др.
На территории Горного Алтая произрастает 4 реликтовых вида рода подмаренник – Galium L. (сем. Rubiaceae): G. krylovii (IIjin) Pobed., G. trifidum L., G. paradoxum Maxim., G. triflorum Michxaux. Подавляющее большинство видов сконцентрировано в Северном и Центральном Алтае.
Реликтовые виды – это остатки древней флоры, когда-то широко распространенные. Такие виды могут быть широко распространены, но чаще они имеют реликтовый ареал, занимаемый видом с момента его вхождения в состав древней флоры [1].
Подмаренник Крылова – G. krylovii IIjin (Cruciata krylovii (IIjin) Pobed.) – многолетнее корневищное растение 10–40 см высоты, стебли высокие или при основании восходящие, лишь по ребрам рассеянно опушены отстоящими волосками (до 0,5 мм), в узлах с кольцом волосков. Листья по 4 в мутовке, с 3 жилками, 10–16 мм длиной и 3–6 мм шириной, от ланцетно-яйцевидные до эллиптических, туповатые, с верхней стороны по краям, а снизу по жилкам усажены короткими (0,3 мм), серповидно изогнутыми, направленными к верхушке щетинками. Соцветия пазушные, малоцветковые, развиваются в средней части растения. Цветоносы и цветоножки голые. Венчик зеленовато-желтый, колесовидный, до 2,5 мм диам., с 4 лопастями. Завязи и плоды голые. Мерикарпий часто 1 вследствие недоразвития второго, шаровидно-почковидный, 2–3,5 мм длиной [5].
Произрастает в Майминском, Турочакском, Чойском, Чемальском, Шебалинском районах, а также встречается в высокогорьях на многих хребтах: Семинском (во многих пунктах по долинам рр. Сема, Анос, Еланда, Каспа, Урсул и ее притоки рр. Курата, Теньга, Табатой), Катунский (в долинах рр. Ак-кем, Кучерла, Кураган, Мульта, Зайчиха), Теректинском (в долинах рр. Каракол, Тюнгур), Курайском (в долине р. Мены – притоках р. Чуи и р. Кубадру – левый приток р. Башкаус), довольно часто на территории Алтайского государственного заповедника [6]. Растет в лесном поясе в темнохвойных пихтовых, еловых и лиственничных лесах, на лесных лугах и в зарослях кустарников на высоте до 2000 м над уровнем моря. Цветет в июне-июле, плодоносит в августе.
Принадлежит к третичному неморальному комплексу растений Сибири. М.М. Ильиным (1941) отнесен к числу третичных широколиственных реликтовых видов, свойственных черневой тайге [7,11].
Подмаренник трехцветковый – G. triflorum Michxaux.– многолетнее растение. Стебли высокие 25–60 см слабые, восходящие или опирающиеся на другие растения, простые, реже разветвленные, голые или рассеянно опушенные отстоящими волосками. Листья короткочерешковые, по 5–6 в мутовках, с 1 жилкой, 13–32 мм длиной и 5–12 мм шириной, от продолговато - ланцетных до эллиптических с коротко остисто-заостренным концом, с верхней стороны по краю усажены прямыми и согнутыми, направленными к верхушке щетинками или голые, с нижней по жилке – обращенными к основанию крючковидными шипиками. Соцветия пазушные, 2, 3, 4-цветковые полузонтики, расположены в средней и верхней части растения. Цветоносы голые, равны или длиннее листьев, из пазух которых выходят и при плодах достигают 40 мм, цветоножки при плодах до 10 мм, голые. Венчик желтовато-зеленоватый, колесовидный, около 3,5 мм диаметром, с 4 длинно заостренными лопастями. Завязи и плоды усажены длинными прямыми щетинками с крючком на конце. Мерикарпии почковидные, около 1,5 мм длиной [5]. Растет в тенистых местах под скалами, в хвойных и смешанных лесах, на лугах. Цветет в июле – августе [9]. Подмаренник трехцветковый, редкий вид флоры Сибири, реликт третичных широколиственных лесов [10].
Произрастает на Семинском хребте (в истоках р. Семы – в долине р. Сарлык), в долине р. Кыги – притоке Телецкого озера, на Курайском хребте (в долине р. Мены и на г. Белькенек) [6].
Подмаренник трехраздельный, п. трехнадрезанный – G. trifidum L. – G. ruprechtii Pobed. – многолетнее растение 5-30 см высотой, простые или разветвленные, полегающиеся или цепляющиеся за другие растения, усаженные по ребрам назад обращенными крючковидными шипиками. Листья по 4 в мутовке, с 1 жилкой, 5–15 мм длиной, 1–3 мм шириной, обратноланцетные, тупые, с верхней стороны усажены мелкими прямыми, направленными к верхушке щетинками или голые, с нижней – по краю и жилке – обращенными основанию крючковидными шипиками. Цветоножки могут быть густо усажены обращенными назад крючковидными шипиками, при плодах изогнутые, 8–10 (13) мм длиной. Венчик белый, колесовидный, около 1–1,5 мм диаметром, 3 (редко 4) лопастями. Завязи и плоды голые. Мерикарпии шаровидные, раздвинутые, около 1,2 мм длиной. Цветет в июне-июле [5].
Растет (1000-2100) на тенистых каменистых склонах, скалах, засоленных низинных лугах, по болотам, заболоченным лесам, по берегам ручьев. Редкий вид в Республики Алтай. Отмечен в ущелье Ачелман- приток р. Чулышман, в Узун-Oюкском районе Алтайского государственного заповедника, в долине р. Чуи в окрестностях с. Кош-Агач и устье р. Чаган-Узун [6].
Подмаренник удивительный – G. paradoxum Maxim. – многолетнее растение. Корневище нитевидное, ползучее. Стебли невысокие (10-20 см), простые, восходящие или полегающие, тонкие, слабые, голые, 4 - гранные. Листья по 4 в мутовке (средние), супротивные, с 1 ясно различимой центральной жилкой, одна пара крупнее другой, накрест лежащей, яйцевидные нежные, тонкие, рассеянно щетинисто-волосистые, нижние более мелкие. Соцветия – верхушечные малоцветковые полузонтики. Цветоносы и цветоножки голые, при плодах удлиняющиеся. Венчик белый, колесовидный или колокольчатый, но без выраженной трубки, до 2,5 мм диаметром, с 4 лопастями. Завязи и плоды покрыты густыми, длинными крючковатыми щетинками. Мерикарпии почковидные, около 1,5 мм длиной [5,8].
Растет в темнохвойных, березовых, осиновых, смешанных лесах, на крупноглыбовых осыпях. Произрастает на Семинском хребте (в истоках р. Семы – в долине р. Сарлык), в долине р. Кыги – притоке Телецкого озера, на Курайском хребте (в долине р. Мены и на г. Белькенек) [6].
G. paradoxum Maxim.– реликт третичных широколиственных лесов. Занесен в Красную книгу РСФСР [11].
Таким образом, исследованные виды рода подмаренник: G. krylovii (IIjin) Pobed., G. paradoxum Maxim., G. triflorum Michxaux являются реликтами третичных широколиственных лесов, а G. trifidum L. – редким видом, поэтому их необходимо изучать и способствовать сохранению во флоре Сибири.
Произрастают данные виды в темнохвойных, березовых, осиновых, смешанных лесах, на крупноглыбовых осыпях, тенистых местах под скалами, на лугах и др. местах. G. krylovii (IIjin) Pobed. и G. paradoxum Maxim. в современных условиях прекрасно приспособлены к условиям их обитания, обладают относительно широкой амплитудой и являются доминантами в некоторых растительных сообществах.
Литература
1. Вульф Е.В. Введение в историческую географию растений. М.–Л.: Сельхозгиз. – 1933. – 415 с.
2 Ревушкин А.С. Высокогорная флора Алтая. Томск: Изд. ТГУ. – 1988. – 320 с.
3. Ильин М.М. Реликтовые элементы широколиственных лесов во флоре Сибири и их возможное происхождение // Тезисы совещания: Проблемы реликтов во флоре СССР. М.-Л.: АН СССР. – 1938. – Вып. 2.– С. 26-31.
4. Красноборов И.М. Третичные реликты во флоре Тувинской АССР Растительный покров бассейна Верхнего Енисея. Новосибирск: Наука. – 1977. – С. 4 -14.
5. Флора Сибири. Т. 12: Solanaceae – Lobeliaceae / сост. А.В. Положий, С.Н. Выдрина, В.И. Курбатский и др. – в 14 томах. – Новосибирск: Наука. Сиб. издательская фирма РАН. – 1996. – 208 с.
6. Ильин В.В, Федоткина Н.В. Сосудистые растения Республики Алтай: аннотированный конспект флоры / Монография. – Горно – Алтайск: РИО ГАГУ. – 2008. – 290 с.
7. Толмачев А.И. Введение в географию растений. Л.: Изд. ЛГУ. – 1974. – 244 с.
8. Редкие и исчезающие растения Сибири. – Новосибирск: Наука. – 1980. – 224 с.
9. Красная книга Республики Алтай (растения). Редкие и находящиеся под угрозой исчезновения виды растений / А.Г. Манеев, И.Н. Пшеничная, Н.В. Федоткина и др. – Новосибирск. – 1996. – 131 с.
10. Семенова Г.П. Редкие и исчезающие виды флоры Сибири: биология, охрана / Г.П. Семенова; Рос. Акад. наук, Сиб. отделение, Центральный сибирский ботанический сад. – Новосибирск: Академическое изд-во «Гео». – 2007. – 408 с.
11. Положий А.В. Реликты третичных лесов во флоре Сибири истории // Известия АН СССР, СО. Сер. биол. Наук. – 1985.
Л. Ситникова
ЖИЗНЕННАЯ ФОРМА И БИОЛОГИЧЕСКАЯ ХАРАКТЕРИСТИКА
CYPRIPEDIUM MACRANTON SW.
Семейство Орхидных (Orchidaceae Lindl.) одно из самых крупных среди однодольных растений. Оно насчитывает около 750 родов и 25 000 видов. Представители семейства очень многообразны по строению и окраске цветков и модификациям вегетативных органов.
По мнению большинства специалистов, орхидные в настоящий момент находятся в стадии активной эволюции. В основном это утверждение касается генеративной сферы растений [1]. Отмечено значительное число межродовых и межвидовых гибридов. Вегетативные органы, обеспечивающие существование растения в тех или иных климатических, эдафических и фитоценатических условиях, остаются более консервативными и отражают в своем строении приспособления к произрастанию именно в тех типах местообитания, в которых проходила эволюция вида. Под влиянием сезонного климата ведущую роль играют видоизменения запасающих органов, способствующие переживанию периодов, неблагоприятных для роста. Например, расположение почек возобновления по отношению к субстрату легло в основу одной из наиболее распространенных классификаций жизненных форм по Раункеньеру. Впоследствии предлагались эколого-морфологические классификации жизненных форм, построенные на основании форм роста, продолжительности жизни, ритма сезонного возобновления, характера питания и т.д. Морфологическое строение побегов системы как основа для выделения жизненных форм использовано в системе И.Г. Серебрякова [7], которая получила в нашей стране широкое распространение. Фитоценотические подходы к классификации биоморф учитывают положение и роль вида в ценозе. Принципы линейной системы жизненных форм, позволяющие достаточно полно и разносторонне охарактеризовать отдельные виды как самостоятельные жизненные формы, использованы в работах В.Н. Голубева, О.В. Смирновой [2, 8].
Башмачок крупноцветковый ( Cypripedium macranton Sw.) растет в лиственничных, сосновых, березовых, смешанных лесах, на лесных сыроватых лугах, среди кустарников, по берегам рек, иногда образует значительные скопления. Он до того прекрасен, что и впрямь может быть посвящен богине любви и красоты. Его волнистые, словно летящие темно-пурпурные лепестки легки и изящны. Губа у башмачка вздута и похожа на переднюю часть атласной туфельки. В народе, поэтому назвали его «Марьин башмачок», «кукушкин башмачок», «богородицыны сапожки». Любопытно, что в Америке этот цветок называют «мокасинами», а в Англии – «дамскими туфельками». А ботаники дали этому растению имя «кипридии (венерин) башмачок». Венера особенно почиталась на острове Кипр, где называлась Кипридой [9].
Вид башмачок крупноцветковый развивается довольно медленно. От прорастания до первого цветения проходит 15 – 17 лет. Морфогенез побегов изучен хорошо. Зачатки внучатой почки обнаруживаются в дочерней почке возобновления в июне, то есть за 2 года до формирования надземного побега. Генеративная сфера в дочерней почке закладывается уже к концу вегетационного сезона [6].
Популяции башмачка в природе бывают очень многочисленными (до нескольких сотен особей). Возрастной спектр правосторонний с преобладанием взрослых вегетативных и генеративных растений (0:8:44:48). Как численность отдельных возрастных групп, так и их соотношения от года к году могут значительно меняться. В отдельные годы максимум численности может смещаться на взрослую вегетативную и даже имматурную группу. Анализ популяций башмачка в разных частях ареала показал, что климатические изменения существенно не влияют на возрастную структуру [4].
Cypripedium macranton Sw. по жизненной форме относится к корневищным орхидным. Он имеет вегетативные побеги, неполный жизненный цикл развития, генеративные побеги моноциклические. Этот вид характеризуется интенсивным вегетативным размножением (до 26% особей в популяции), что ведет к образованию клонов. Вегетативное потомство относится к тому же возрастному состоянию, что и материнское растение, либо неглубоко омоложено. Ювенильные и имматурные растения вегетативно не размножаются [5].
Наши полевые исследования за Cypripedium macranton Sw. ведутся с 2005 года на территории агробиостанции ГАГУ. В отдел редких и исчезающих растений нами была привезена популяция этого вида из с. Каракокша Чойского района. Сезонные изменения проводились на опытном участке. Цветение башмачка крупноцветкового происходит в мае – июне и длится 2 – 3 недели. Зрелые семена характеризуются глубоким покоем. Плодоношение наступает в августе. Плоды образуются нерегулярно. Колебания в плодообразовании в большинстве случаев объясняются не климатическими причинами, а недостаточным опылением. Виды рода Cypripedium являются слабоспециализированными энтомофилами. Опыление осуществляется нерегулярно мелкими насекомыми из разных отрядов.
Наибольшую угрозу рассматриваемому виду представляют рекреация вблизи городов, дачных участков и др., выкапывание и пересадка в сады взрослых растений, сбор букетов, вытаптывание и повреждение растений. Высокая декоративность цветков Cypripedium macranton Sw. делает их особенно привлекательными для людей. Необходима тщательная охрана местообитаний башмачков, а также пропаганда природоохранных идей среди местного населения. Кроме того, в последние годы башмачки становятся объектом коммерческой торговли и вывозятся за границу. В труднодоступных регионах еще сохраняются крупные популяции этих видов. Вид Cypripedium macranton Sw. занесен в красную книгу РСФСР [3].
Литература
1. Аверьянов Л.В. Конспект рода Dactylorhiza Neck. Ex Nevski (Orchidaceae), 1 // Новости систематики высш. растений. – Л., 1990. – Т. 27. – С. 32 – 62.
2. Голубев В.Н. Принцип построения и содержание линейной системы жизненных форм покрытосеменных растений // Бюл. МОИП. Отд. Биол. – 1972. – Т. 77, вып. 6. – С. 72 – 80.
3. Красная книга РСФСР (растения). – М.: Росагропромиздат, 1988. – 476 с.
4. Куликов П.В. Экология и репродуктивные особенности редких орхидных Урала: Автореф. дис… канд. биол. наук. – Екатеринбург, 1995. – 24 с.
5. Кулль Т.В. Вегетативное размножение Cypripedium сalceolus L. в естественных условиях // Охрана и культивирование орхидей. – М., 1987. – С. 13 – 14.
6. Полынова Н.А., Утёмова Л.Д. Биологические особенности видов рода Венерин башмачок в условиях Хакасской автономной области // Исчезающие и редкие растения и животные Алтайского края и пробл. их охраны. – Барнаул, 1982. – С. 78 – 81.
7. Серебряков И.Г. Жизненные формы высших растений // Полевая геоботаника – М.; Л., 1964. – Т. 3. – С. 146 – 205.
8. Смирнова О.В. Структура травянистого покрова широколиственных лесов. – М.: Наука, 1987. – 205 с.
9. Татаренко И.В. Орхидные России: жизненные формы, биология, вопросы охраны. – М.: Аргус, 1996. – 207 с.
Т.В. Тельминова
ОНТОМОРФОГЕНЕЗ И СЕЗОННЫЙ РИТМ РАЗВИТИЯ
CENTAUREA SCABIOSA l.
Василек шероховатый – травянистый длинностержнекорневой факультативно корнеотпрысковый многоглавый поликарпик с симподиальной системой полурозеточных побегов [4].
У семян василька слабо развита семенная кожура, они плотно прилегают к околоплоднику. В зародыше содержится много масла. Семена могут прорастать без периода покоя, но в год созревания прорастают плохо, дружные всходы появляются лишь на следующий год [1].
Прорастание семян надземное. Семядоли 12-15 мм длиной и 5-7 мм шириной продолговато-обратнояйцевидные или продолговатые, постепенно суженные в короткие широкие черешки, цельнокрайные, с тупой верхушкой и едва заметной средней жилкой, темно-зеленые, слегка мясистые, располагаются почти горизонтально [2].
Из верхушечной почки вырастает розеточный побег, первые листья которого (их бывает 2-3) цельные, продолговатые, с острой верхушкой, почти цельнокрайние. Длина их 2-4 (5-8) см, ширина 5-10 мм. Главный корень несколько более тонкий, чем гипокотиль, начинает ветвиться в базальной части уже у трехдневных проростков. С появлением новых листьев корневая система начинает нарастать, возникают корни третьего порядка.
К середине или к концу вегетационного периода семядоли засыхают, вырастают новые листья, гипокотиль укорачивается и утолщается, растение переходит в ювенильное состояние. Длинночерешковые продолговатые листья ювенильных растений направлены косо вверх и могут достигать длины 19 см, но средняя их длина 8 см. В пазухах нижних листьев обнаруживаются открытые почки возобновления, которые оказываются у поверхности почвы вследствие контрактильной деятельности корней, а также за счет укорачивания гипокотиля [3].
Корневая система проникает вглубь более чем на 30 см. Главный корень обильно ветвится в базальной части, в поверхностном слое почвы. Во время перезимовки у ювенильных растений могут сохранятся основания листьев, но они выходят из под снега желтоватыми с фиолетовым оттенком от антоциана. На втором году жизни из верхушечной почки вырастает розеточный побег опять с цельными листьями, но больших размеров. Их средняя длина 14,5 см, максимальная – 22 см, ширина до 2 см, они заметно зазубрены по краю, в розеточном побеге их 4-5. От прошлогодних листьев остаются отмацерированные сосудисто-волокнистые пучки на очень коротком вертикальном корневище [1, 3].
На третьем году жизни, очень редко на втором, у василька появляются листья с первыми признаками рассечения, т. е намечается переход к перистораздельному, а затем перисторассеченному листу взрослых особей. На одной стороне листа или с двух сторон в нижней части появляется по 3-4 крупных зубца или боковых долей, лист становится лировидным. Средняя длина листьев 19 см (до 28), ширина 3-4 см. Растения с такими листьями относятся к имматурному возрастному состоянию. Нередко в розетках имматурных особей нижние листья бывают цельными как у ювенильных. В других случаях наряду с листьями имматурного типа развиваются листья, сходные с листьями взрослых вегетативных особей, т.е. перисторассеченные, но с более крупным верхушечным сегментом. Корневая система имматурных растений углубляется до150- 185 см. Она максимально ветвится в поверхостном слое почвы (до 20см). Среди мелких боковых корней выделяются более крупные горизонтально отходящие корни. Толщина главного корня у основания 3-7 мм, он особо линяет [1,3].
Следующую возрастную группу составляют взрослые вегетативные растения. Их розеточные побеги имеют листья перисторассеченные или двояко-перисторассеченные. И. Г. Серебряков [4] отмечает дивергентный тип формирования листовой пластинки у василька шероховатого. Длина листьев может быть до 38 см, в среднем 25 см. Корневая система углубляется более чем на 2 м. Это возрастное состояние может в онтогенезе выпадать, если василек растет на рыхлой почве. Тогда особи, перезимовав, могут переходить к цветению.
На рыхлой почве на третий год, а в не нарушенных луговых ценозах на пятый – шестой год растения обычно переходят в генеративное состояние. На верхушке розеточного побега, нараставшего в виргинильный период моноподиально, закладывается цветочная почка, из которой развивается генеративный побег первого порядка. В конце вегетационного периода он отмирает почти полностью, остается лишь небольшой пенек. На следующий год происходит перевершинивание одного или двух генеративных побега. С этого момента начинается образование многоглавого каудекса.
Растения, цветущие первый и второй раз, составляют группу молодых генеративных особей. Цветоносные побеги их обычно не высокие, в среднем 53 см (32-75 см), слабо ветвятся и несут всего лишь несколько корзинок. Каудекс развит слабо, главный корень еще не толстый и не затронут процессами разрушения. У молодых генеративных особей может наступить перерыв в цветении.
В группу средневозрастных генеративных растений входят особи, как правило, с хорошо развитым, разветвленным, до 20 см в поперечнике, начавшем разрушаться в центре каудексом, от которого отходит несколько генеративных побегов, достигающих в высоту 60-80, иногда 120-150 см. Корневая система, углубляющаяся до 2,5 м, имеет крупные боковые скелетные корни, отходящие от главного вместе с тонкими [1,3].
У старых генеративных растений василька (Centaurea scabiosa), возраст которых 20-30 лет, отмирают некоторые главы каудекса, внутри главного корня, достигающего толщины 5-6,5 см, возникает крупная полость, заполняющаяся почвой, куда проникают корневища соседних растений, заползают дождевые черви и другие животные, начинается партикуляция. Материнская особь может распадаться на две или несколько партикул, несущих генеративные побеги. Некоторое время они могут существовать самостоятельно благодаря тому, что каждая партикула снабжена одним или несколькими крупными корнями, а также мелкими придаточными корнями. Однако все партикулы дуплисты, сильно разрушаются ткани корня как внутри, так и снаружи, поэтому они быстро стареют и переходят в сенильное состояние. В сенильное состояние могут переходить также и непартикулирующие особи [1,3].
У сенильных растений сильно разрушен каудекс, развиваются только вегетативные побеги с двояко-перисторассеченными листьями, более короткими, чем у взрослых вегетативных особей.
В окрестностях города Горно-Алтайска всходы появляются в массе весной, но могут появляться в конце лета (после обильных теплых дождей) и осенью. Переход в ювенильное состояние совершается к концу вегетационного периода или после перезимовки. Молодые растения василька зимнезеленые, в то время как взрослые-летнезеленые. Вегетация начинается во вторую декаду после таяния снега, т. е обычно в начале или в конце апреля. Почки открытые. Листья весеннее-летней генерации продолжают появляться еще и в июне. Цветочные почки закладываются в год цветения. Цветение обычно начинается в начале июля и длится до сентября – октября. Из-за длительного цветения плодоношение тоже растянуто. На одном растении могут быть соцветия одновременно со зрелыми или созревающими плодами, цветками и бутонами. Начало плодоношения – в конце июля – начале августа, а массовое плодоношение наступает во второй декаде августа. В поздно цветущих корзинках плоды не вызревают. К концу плодоношения стеблевые листья засыхают, остается живым стебель, Который засыхает после обсеменения.
Centaurea scabiosa – гемикриптофит
Литература
1. Былова А.М. Вопросы морфогенеза цветковых растений и строение их популяций. – М.: Наука
2. Былова А. М. Регенерация Centaurea scabiosa L. // Биологические науки, № 1
3. Еленевский А. Г. Ботника: Систематика высших, или наземных, растений: Учебник для высших педагогических учебных заведений /А. Г. Еленевский, М. П. Сословьева, В. Н. Тихомиров.-3-е изд., испр. И доп. – М.: Издательский центр «Академия», 2004. - 432 с.
4. Серебряков И.Г. Экологическая морфология растений. Жизненные формы покрытосеменных и хвойных /И.Г. Серебряков. М.: Высшая школа, 1962. - 378 с.
А.А. Тымтышева
МОРФОЛОГИЧЕСКИЕ ПОКАЗАТЕЛИ НОВОРОЖДЕННЫХ ДЕТЕЙ ВЫСОКОГОРНОЙ И НИЗКОГОРНОЙ ЗОН ГОРНОГО АЛТАЯ
Последнее десятилетие XX века характеризовалось в РФ значительной убылью населения из-за резкого снижения рождаемости и высокого уровня общей смертности. При этом более трети детей рождались с отклонениями в состоянии здоровья, общая заболеваемость новорожденных почти в два раза превышала уровень, отмеченный в предыдущем десятилетии.
Изучение морфологического развития новорожденного имеет значение не только для оценки настоящего состояния организма, но и для прогнозирования уровня психического и физического здоровья, антропометрических признаков в последующие периоды онтогенеза, что в значительной степени увеличивает научно-практический интерес исследования.
Уровень морфологического состояния детей тесно взаимосвязан с социально-экономическими, климатогеографическими, региональными, эколого-гигиеническими и другими условиями.
Целью данного исследования явилось изучение морфологических показателей новорожденных детей высокогорной и низкогорной зон Горного Алтая в период с 2002 по 2004 гг. Период исследования выбран в связи с тем, что в Горном Алтае произошло Алтайское (Чуйское) землетрясение 27 сентября 2003 г. и одной из задач явилось рассмотрение вопроса о его влиянии на морфологическое состояние детей.
Сбор первичного материала проводился в высокогорном Кош-Агачском районе (где находился эпицентр землетрясения) и в низкогорном Чемальском районе Республики Алтай. Материалом работы послужили медицинские карты новорожденных детей. Проанализировано 314 медицинских карт новорожденных детей, из них 232 высокогорной зоны (103 мальчика, 129 девочек) и 82 низкогорной зоны (38 мальчиков, 44 девочки).
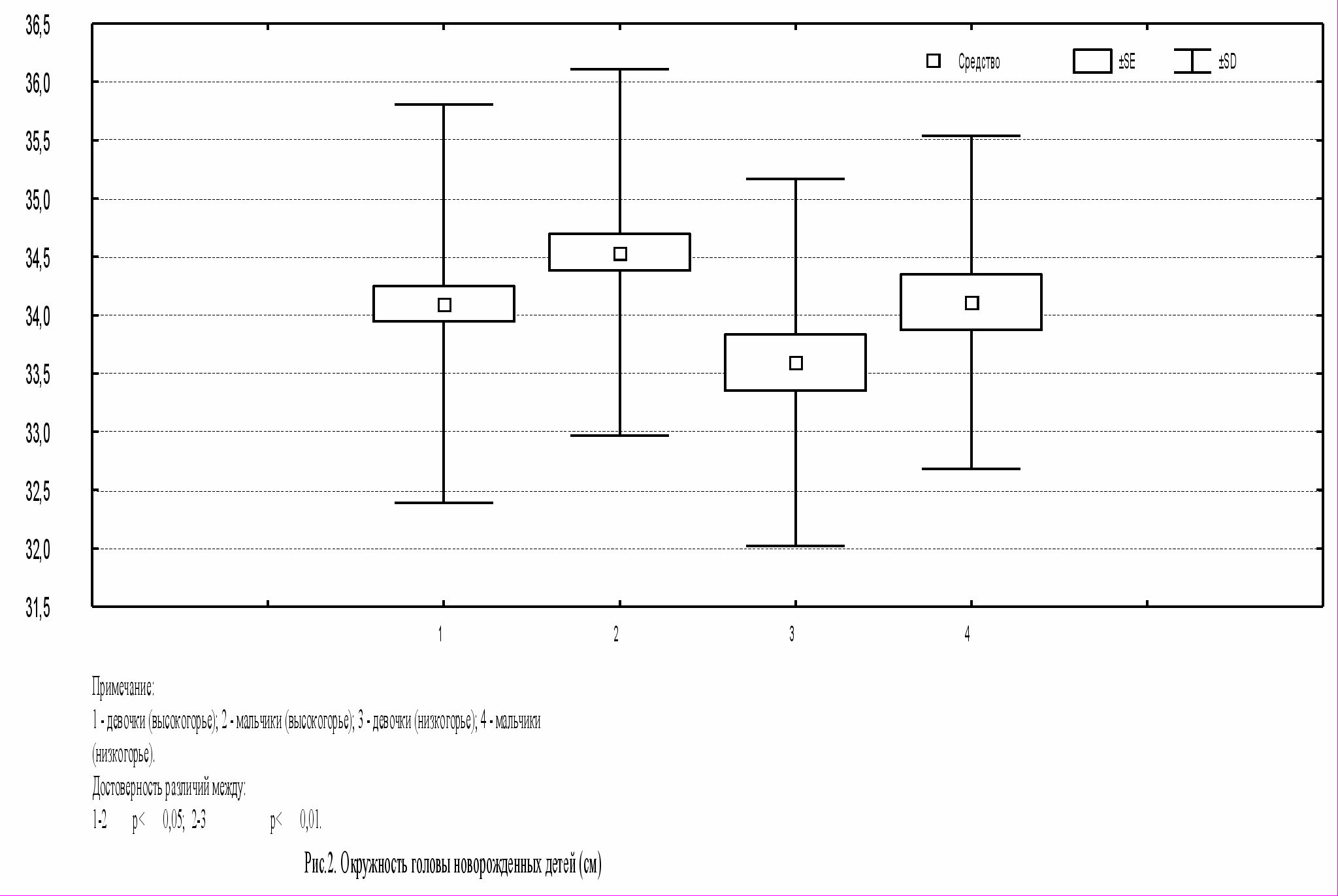
В результате исследования было выявлено, что по длине тела новорожденные высокогорной зоны имеют значения ниже по сравнению с детьми низкогорной зоны (p<0,05; p<0,01; рис.1). По массе тела среди исследуемых групп достоверных различий не выявлено, показатель находился в пределах 3200 – 3350 г. Средние значения окружности грудной клетки составили от 32,8 см до 33,5 см, по окружности головы – 33,5 – 34,5 см, однако не показали достоверных различий в изучаемых группах.
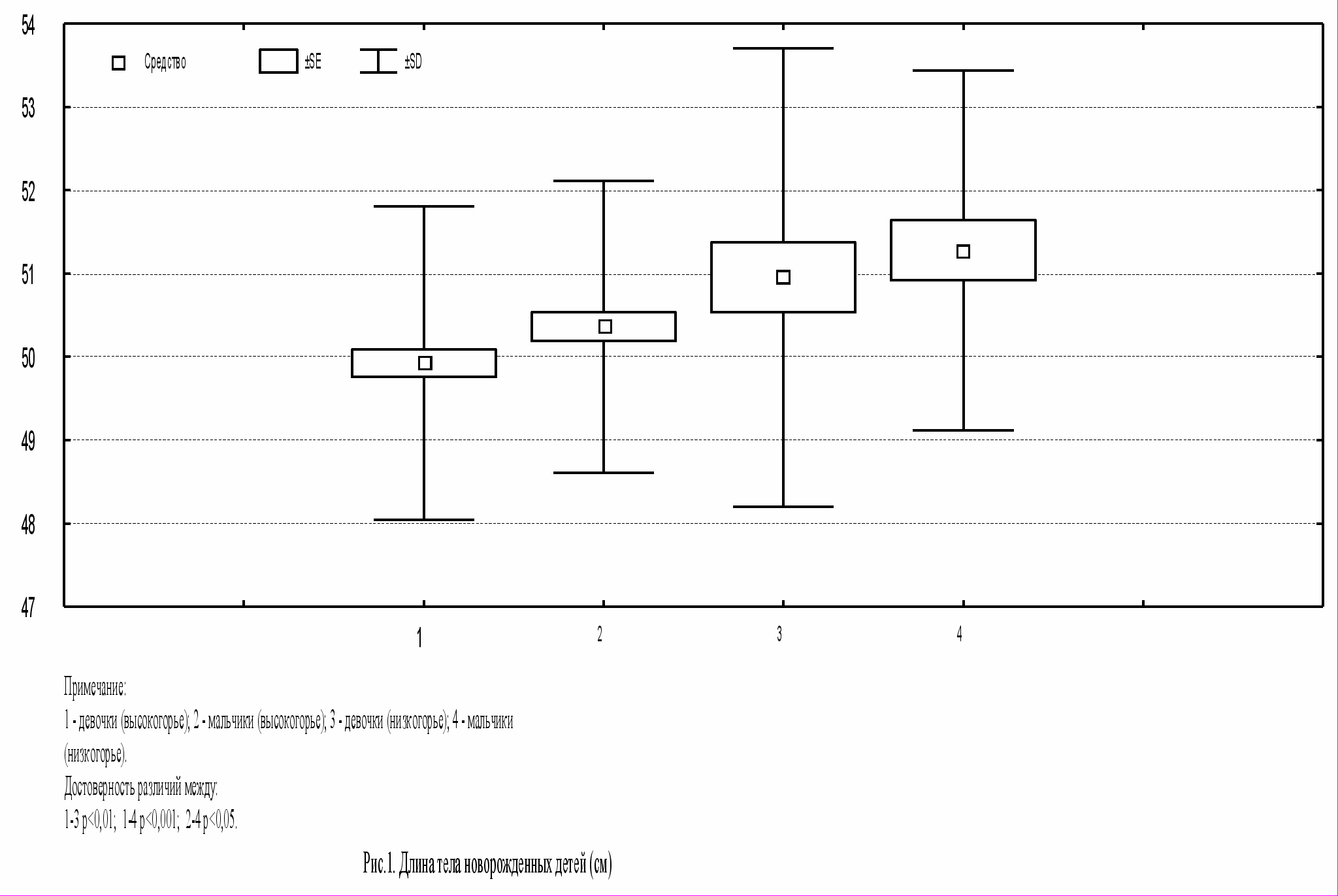
В этой связи уместно предположить, что морфологическое состояние детей зависит от климатогеографических условий.
Для изучения различий по изучаемым антропометрическим признакам между мальчиками и девочками мы использовали абсолютные показатели. По средним значениям массы и длины тела, окружности грудной клетки мальчики больше девочек. Различия по окружности головы между мальчиками и девочками (рис.2) статистически достоверны (p<0,05; p<0,01). Наши результаты согласуются с данными литературы, что средние значения новорожденных мальчиков больше, чем новорожденных девочек.
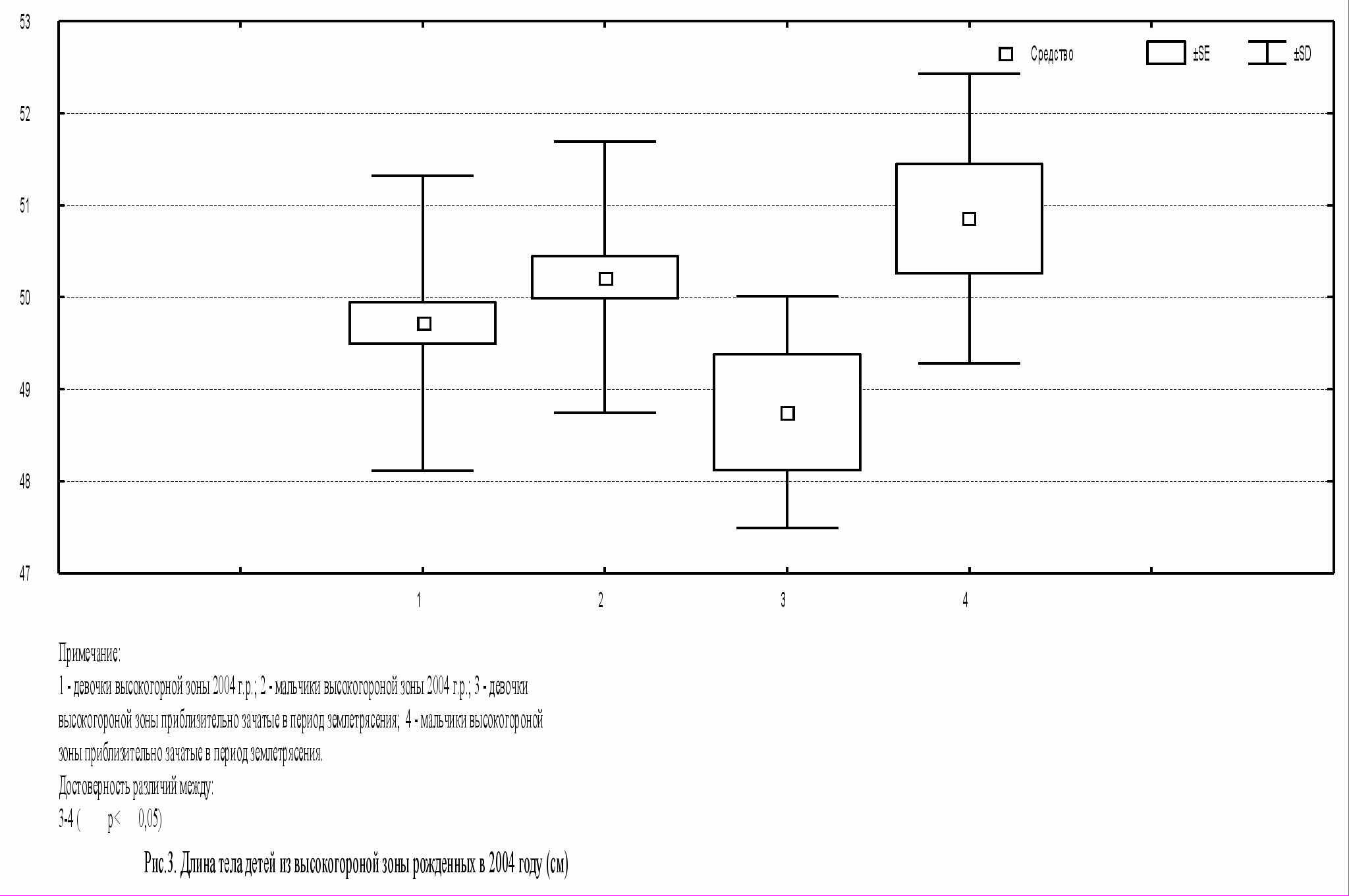
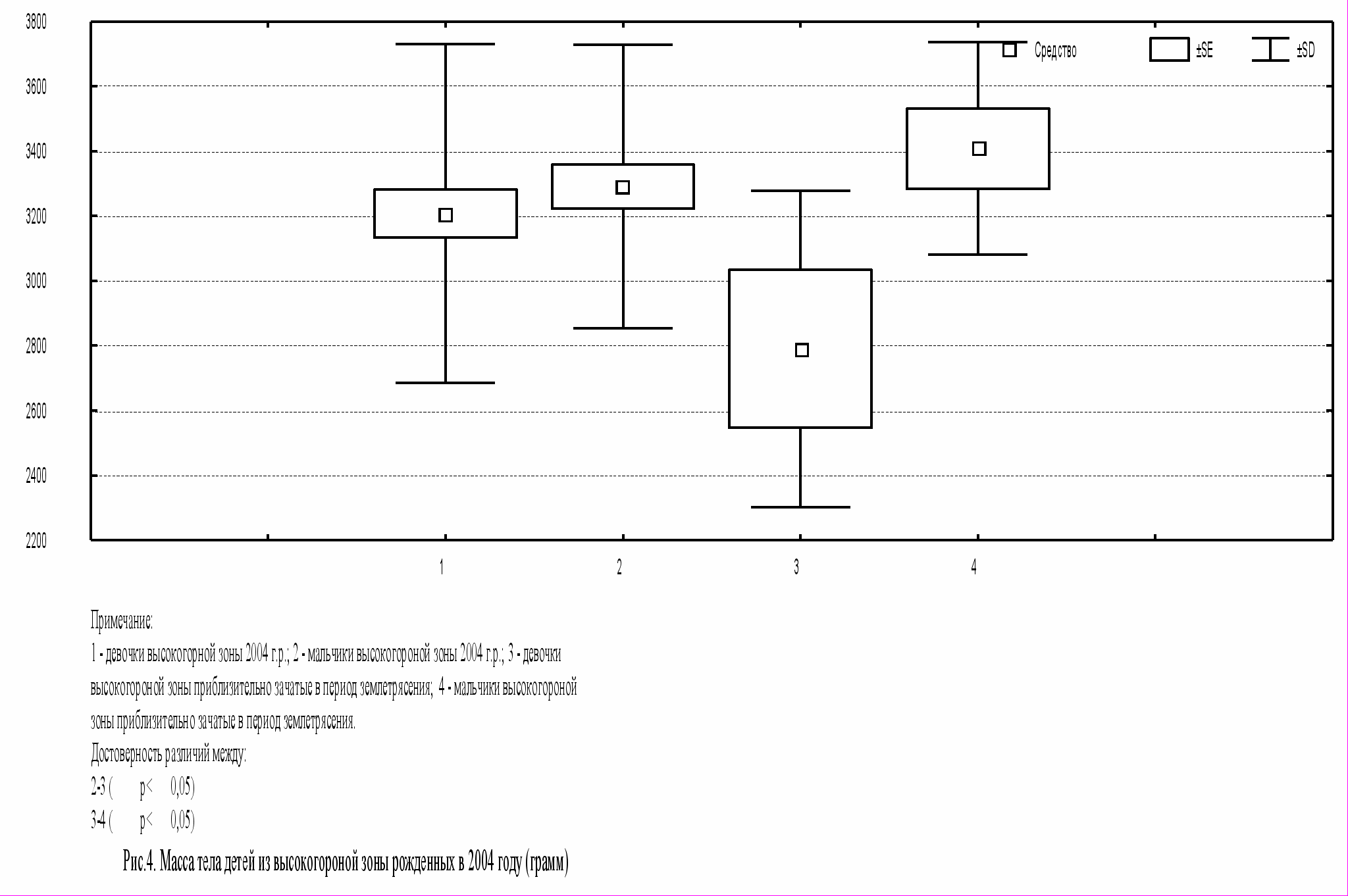
При рассмотрении вопроса о возможном влиянии землетрясения на тотальные размеры тела новорожденных исследуемые группы детей высокогорной зоны были поделены на две группы. Первую группу составили дети, зачатые до и после землетрясения, во вторую вошли новорожденные, приблизительно зачатые в период землетрясения. В результате сравнения была выявлена тенденция к снижению тотальных размеров тела у девочек 2 группы, но к увеличению новорожденных мальчиков 2 группы.
При этом статистически значимые данные были получены только по длине и массе тела среди детей второй группы. Как видно из рисунков 3-4 новорожденные девочки имеют длину тела на 2 см (p<0,05), а массу тела на 600 г (p<0,05) меньше, чем новорожденные мальчики. Таким образом, вероятно новорожденные девочки оказались более чувствительны к изменению сейсмологических условий.
Литература
1. Баранов, А.А. Здоровье детей России / А.А. Баранов. – М., 1999.
2. Дмитриев, Д.А. Значение размеров тела при рождении для прогнозирования состояния здоровья в последующие периоды онтогенеза / Д.А. Дмитриев, А.Д. Дмитриев, М.Н. Ташкова // Профилактика заболеваний и укрепления здоровья. - 2003. - №2.-С.19-25.