
Общая биология уровни организации и основные признаки живого вещества уровни организации живого вещества молекулярный
| Вид материала | Документы |
- Перечень вопросов для курсового экзамена по биологии уровни организации живого. Человек, 96.87kb.
- Чебышев Н. В. Биология: учебник для студентов сред. Проф. Учебных заведений, 197.24kb.
- «биология», 388.53kb.
- Экзаменационные вопросы по биологии биология: определение, современный этап развития, 123.72kb.
- Оглавление лекция №1, 787.83kb.
- Основные вопросы программы по биологии для студентов 1 курса заочного обучения по специальности, 55.73kb.
- Основные вопросы программы по биологии для студентов 1 курса заочного обучения волгоградской, 648.74kb.
- Размножение и индивидуальное развитие организмов, 44.8kb.
- Урок биологии-9 класс. Тема «Многообразие живого мира. Основные свойства», 52.53kb.
- Учебно-тематический план по биологии 9 класс (70 ч.), 98.84kb.
ВЗАИМОДЕЙСТВИЕ ГЕНОВ
I. Плейотропия (множественное действие генов): один ген отвечает за проявление нескольких признаков в фенотипе.
II. Взаимодействие аллельных генов.
Неполное доминирование: в случае гетерозиготы ни один из аллельных генов не подавляет проявление другого гена в фенотипе.
Кодоминантность: один признак определяется не двумя, а тремя (или более) генами, причем два гена доминируют над третьим, но не доминируют друг над другом.
III. Взаимодействие неаллельных генов.
Комплементарность: признак проявляется в случае одновременного присутствия в зиготе двух (или более) опре-деленных неаллельных генов.
Эпистаз: признак определяется двумя (или более) парами аллелей, причем доминантный ген одной пары аллельных генов подавляет проявление доминантного гена другой пары аллельных генов.
Полимерия: один признак определяется двумя парами аллелей, причем доминантный ген одной пары аллелей усили-вается действием второй.
ИЗМЕНЧИВОСТЬ
I. Ненаследственная (фенотипическая, групповая, опре-деленная, модификационная) изменчивость не связана с изменением генотипа.
Норма реакции – это пределы модификационной изменчи-вости признака. У гибридов преимущественно доминируют те признаки, которые в окружающей среде встречают наиболее благоприятные условия для своего развития.
Вариационный ряд – это порядок расположения числа вариант по мере увеличения данного признака (варианта – это определённая величина признака.
Вариационная кривая – это график зависимости частоты встречаемости вариант от величины признака.
Средняя величина признака определяется по формуле:
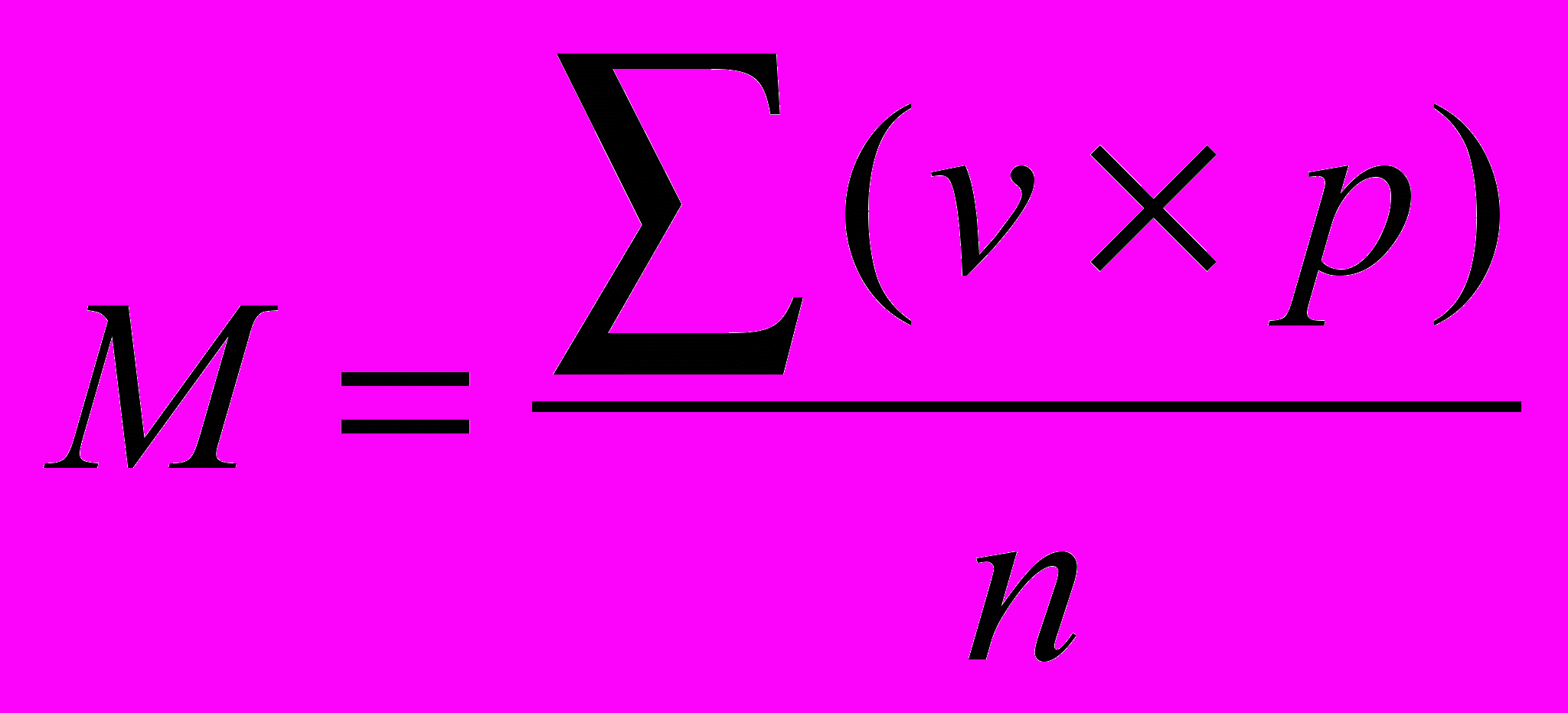 ,
, где M – средняя величина, v – варианта, p – частота встречае-мости вариант, n – общее число вариант вариационного ряда.
II. Наследственная (генотипическая, неопределенная, или индивидуальная) изменчивость обусловлена изменениями генотипа в той или иной степени.
1. Мутационная изменчивость
Генные мутации:
делеция – выпадение пар нуклеотидов из ДНК,
дупликация – повторение пар нуклеотидов,
инсерция – вставка пар нуклеотидов,
инверсия - перестановка нуклеотидов,
замена нуклеотидов.
Геномные мутации
гаплоидия - кратное уменьшение числа хромосом (каждая хромосома представлена в наборе в единичном числе);
полиплоидия - кратное увеличение числа хромосом;
анеуплоидия - изменение числа хромосом за счет добавле-ния или потери отдельных хромосом;
Структурные мутации хромосом:
делеция - потеря того или иного участка хромосом;
дупликация - удвоение отдельного участка хромосом;
инверсия - поворот участка хромосом на 180%.
2. Комбинативная изменчивость проявляется у гибрид-ных потомков в результате всевозможных комбинаций родительских генов как в старых, так и в новых сочетаниях, за счёт неполного сцепления генов в хромосомах или взаимодействия генов в новых комбинациях.
3. Соотносительная изменчивость связана с множествен-ным действием генов.
ЗАКОН ГОМОЛОГИЧЕСКИХ РЯДОВ В НАСЛЕДСТВЕННОЙ ИЗМЕНЧИВОСТИ
Виды и роды, генетически близкие, характеризуются сходными рядами наследственной изменчивости с такой правильностью, что, зная ряд форм в пределах одного вида, можно предвидеть нахождение параллельных форм у других видов и родов. Чем ближе генетически расположены в общей системе роды и виды, тем полнее сходство в рядах их изменчивости. Целые семейства растений в общем характеризуются определенным циклом изменчивости, проходящей через все роды и виды, составляющие семейство.
ЗАКОН ХАРДИ - ВАЙНБЕРГА
В случаи полной изоляции популяции и отсутствии возникновения мутаций соотношение генов в генофонде данной популяции остаётся неизменном из поколения в поколение.
ИСКУССТВЕННЫЙ МУТАГЕНЕЗ
Мутагенез – процесс возникновения мутаций в организме. Может быть спонтанным (самопроизвольным) и индуцированным (вызванным искусственно различными мутагенами).
Естественные мутации, сопровождающиеся проявлением полезных для человека признаков, возникают очень редко. Частота мутаций резко повышается при воздействии мутагенов. К ним относятся некоторые химические вещества, а также ультрафиолетовое и рентгеновское излучение. Эти воздействия нарушают строение молекул ДНК и служат причиной резкого возрастания частоты мутаций. Наряду с вредными мутациями нередко обнаруживаются и полезные, которые используются учеными в селекционной работе. Мутагенез широко применяется в создании штаммов микрооорганизмов. Путем воздействия мутагенами в растениеводстве получают полиплоидные растения, отличаю-щиеся более крупными размерами, высокой урожайностью и более активным синтезом органических веществ.
ОСНОВЫ СЕЛЕКЦИИ
ЗАДАЧИ СОВРЕМЕННОЙ СЕЛЕКЦИИ
Селекция разрабатывает теорию и методы выведения новых и улучшения уже имеющихся пород животных, сортов растений и штаммов микроорганизмов. Основами селекции служат: теория искусственного отбора, генетика, молекуляр-ная биология, экология, биогеография. Большой вклад в развитие селекции в нашей стране внесли такие известные учёные как: Н.И. Вавилов, И.В. Мичурин, П. П. Лукьяненко, В. С. Пустовойт, В. Н. Ремесло, М. Ф. Иванов и др.
Породой животных или сортом растений называют такую совокупность особей, искусственно созданную человеком, которая характеризуется определенными наследственными особенностями: продуктивностью, морфологическими и физиологическими признаками.
ОСНОВНЫЕ МЕТОДЫ СЕЛЕКЦИИ
Отбор
Массовый отбор сводится к выделению из исходного материала целой группы особей, которые обладают желательными для селекционера признаками. В растение-водстве массовый отбор применяется по отношению к перекрестно опыляющимся растениям. При повторном посеве вновь отбирают растения с заданными признаками. Сорт, получаемый этим способом, не является генетически однородным, и отбор приходится повторять.
Индивидуальный отбор сводится к выделению отдельных особей с определёнными признаками и получению от них потомства, он может быть однократным и повторяющимся.
В селекции животных применяется жесткий отбор по таким признакам как: выносливость, экстерьер, продуктив-ность и др. В растениеводстве – применяется в отношении самоопыляющихся растений, выделяются чистые линии, то есть потомство одной самоопыляющейся особи.
Гибридизация:
Неродственная гибридизация (аутбридинг) в селекции животных - скрещивание отдаленных пород, отличающихся контрастными признаками, для получения гетерозиготных популяций и проявления гетерозиса, получается бесплодное потомство; В селекции растений – внутривидовое, межвидо-вое, межродовое скрещивание, ведущее к гетерозису.
Близкородственная гибридизация (инбридинг) – скрещива-ние между близкими родственниками для получения гомозиготных линий с желательными признаками. В селекции растений – самоопыление у перекрестноопыляющихся расте-ний путем искусственного воздействия на них.
Испытания производителей по потомству
В селекции животных – используют метод искусственного осеменения от лучших производителей, качества которых проверяют по многочисленному потомству.
Производитель – самец, обладающий ценными признаками, по которым ведется селекция. Основные племенные качества производителя – экстерьер и продуктивность.
Подбор родительских пар
В селекции животных – по хозяйственно ценным признакам и по экстерьеру; в селекции растений – по месту их происхождения (географически удаленных) или генетически отдаленных (неродственных).
Экспериментальное получение полиплоидов
В селекции растений применяется для получения более продуктивных и урожайных форм.
Самоопыление перекрестноопыляемых растений
Самоопыление ведет к повышению гомозиготности, что способствует закреплению наследственных свойств, но оно приводит к снижению жизнеспособности, уменьшению продуктивности, к вырождению. Несмотря на неблагоприятное влияние самоопыления, у перекрестно-опыляемых растений его часто и успешно применяют в селекции. Обычно сначала выводят гомозиготные линии, у которых закрепляются желательные признаки. Вместе с тем происходит резкое снижение урожайности. Затем проводят перекрестное опыление опыление между разными самоопыляющимися линиями, в результате чего в ряде случаев появляются высокоурожайные гибриды.
При скрещивании разных пород животных или сортов растений, а также при межвидовых скрещиваниях в первом поколении гибридов повышается жизнеспособность и наблюдается мощное развитие. Это явление получило название гибридной силы или гетерозиса. Оно объясняется переходом многих генов в гетерозиготное состояние и взаимодействием благоприятных доминантных генов. При последующих скрещиваниях гибридов между собой гетерозис затухает вследствие выщепления гомозигот.
МЕТОДЫ СЕЛЕКЦИОННОЙ РАБОТЫ И. В. МИЧУРИНА
Биологически отдаленная гибридизация
Межвидовая гибридизация – скрещивание представителей разных видов для получения сортов с нужными свойствами (например, вишня Владимирская + черешня Винклера белая = вишня Краса севера (хороший вкус, зимостойкость));
Межродовая гибридизация – скрещивание представителей разных родов для получения новых форм растений (например, вишня + черемуха = церападус).
Географически отдаленная гибридизация
Скрещивание представителей контрастных природных зон и географически отдаленных регионов производят с целью придать гибриду нужные качества (например, груша дикая уссурийская + Бере рояль (Франция) = Бере зимняя Мичурина). Однако отдаленные гибриды обычно бесплодны. Основной причиной бесплодия у отделенных гибридов обычно является невозможность нормального хода созревания половых клеток; хромосомы обоих родительских видов растений настолько несхожи между собой, что они оказываются неспособными конъюгировать, в результате чего нарушается процесс мейоза.
Отбор
Многократный, жесткий по многим параметрам, таким как: размер, форма, зимостойкость иммунные свойства, качество, вкус, цвет плодов, и их лёжкость (много сортов яблонь с хорошими вкусовыми качествами и высокой урожайностью продвинуто на север).
Метод ментора
Воспитание в гибридном сеянце желательных качеств (усиление доминирования) осуществляется путём прививки привоя на растение-воспитатель, от которого эти качества хотят получить, причём чем ментор старше, мощнее, дольше действует, тем его влияние ощутимей, например: яблоня Китайка (подвой) + гибрид (Китайка + Кандиль-синап) = Кандиль-синап (морозостойкий); Бельфлер-китайка (гибрид-подвой) + Китайка (привой) = Бельфлер-китайка (лёжкий позднеспелый сорт).
Подвой – растение, на которое производится прививка.
Привой – черенок растения или почка, которые прививаются на подвой.
Метод посредника
При отдаленной гибридизации для преодоления нескре-щиваемости различных сортов используют дикий вид растения в качестве посредника, который будет переносить желаемый признак от исходного к конечному гибриду например: дикий монгольский миндаль + дикий персик Давида = миндаль Посредник; Культурный персик + миндаль Посредник = гибридный персик (продвинут на север.
Воздействие условиями среды
При воспитании молодых гибридов обращалось внимание на метод хранения семян, характер и степень питания, воздействие низкими температурами, бедной питанием почвой, частыми пересадками (например, закаливание гибридного сеянца; отбор наиболее выносливых растений).
Смешение пыльцы
Для преодоления межвидовой нескрещиваемости (несовместимости) смешивалась пыльца материнского растения с пыльцой отцовского растения другого вида, материнская пыльца раздражала рыльце, и оно воспринимало чужую оцовскую пыльцу.
ЦЕНТРЫ МНОГООБРАЗИЯ И ПРОИСХОЖДЕНИЯ
КУЛЬТУРНЫХ РАСТЕНИЙ
1. Южноазиатский тропический центр. Тропическая Индия, Индокитай, Южный Китай, острова Юго-Восточной Азии очень богаты культурными растениями (около половины известных видов). Родина риса, сахарного тростника, множества плодовых и овощных культур.
2. Восточноазиатский центр. Центральный и Восточный Китай, Япония, остров Тайвань, Корея - родина сои, нескольких видов проса, множества плодовых и овощных культур. Около 20 % видов культурных растений.
3. Юго-Западноазиатский центр. Малая Азия, Средняя Азия, Иран, Афганистан, Северо-Западная Индия - родина нескольких форм пщеницы, ржи, бобовых, винограда, плодовых. В нем возникло 14% мировой культурной флоры.
4. Средиземноморский центр. Страны, расположенные по берегам Средиземного моря. Около 11% видов культурных растений, в числе которых маслины, многие кормовые растения (клевер, одноцветковая чечевица), многие овощные (капуста) и кормовые культуры.
5. Абиссинский центр. Небольшой район Африканского материка (территория Эфиопии) с очень своеобразной флорой - родина зернового сорго, одного вида бананов, масличного растения нута, ряда особых форм пшеницы и ячменя.
6. Центральноамериканский центр. Южная Мексика –
родина около 900 видов культурных растений: кукурузы, длинноволокнистого хлопчатника, какао, тыквы, фасоли и др..
7. Андийский (Южноамериканский) центр. Включает часть района Андийского горного хребта вдоль западного побережья Южной Америки - родина многих клубненосных растений, в том числе картофеля, некоторых лекарственных растений (кокаиновый куст, хинное дерево и др.).
Центры происхождения культурных растений тесно связаны с районами одомашнивания животных, они получили назва-ние центров доместикации.
СЕЛЕКЦИЯ МИКРООРГАНИЗМОВ
Микроорганизмы интенсивно используются в самых разнообразных технологических процессах. В связи с этим развивается промышленная микробиология и ведется интенсивная селекция новых штаммов микрооргаизмов с повышенной продуктивностью веществ, необходимых человеку. Технологию получения неоходимых человеку продуктов из живых клеток или с их помощью называют биотехнологией. Большое значение в биотехнологии приобретают методы, получившие название клеточной инженерии. Предварительно клетки искусственно выделяют из организма и переносят на специально созданные питательные среды, где они в стерильных условиях продолжают жить и размножаться. Такие клеточные культуры (или культура тканей) могут служить для продукции ценных веществ. Клеточные культуры используют и для гибридизации клеток. Применяя некоторые специальные приемы, можно объединить клетки разного происхождения от организмов, обычная гибридизация которых половым путем невозможна. В биотехнологии широко применяется метод генной (генетической) инженерии, которая представляет собой совокупность методов молекулярной генетики, связанных с искусственным созданием новых комбинаций генетического материала, способного размножаться в клетке-хозяине и синтезировать конечные продукты обмена. Задача этих методов состоит в получении индивидуальных генов и введении их в новое генетическое окружение для создания организмов с новыми, заранее предопределенными признака-ми. Основные этапы создания генетически измененных организмов следующие:
а) получение гена, кодирующего интересующий признак;
б) объединение этого гена с вектором;
Вектор представляет собой плазмиду или вирус (по химическому составу является фрагментом ДНК), в геном которых можно внедрить чужеродную генетическую информацию. Вектор обеспечивает протекание следующих процессов:
а) встраивание рекомбинантной ДНК в генетический аппарат клетки-хозяина;
б) размножение рекомбинантной ДНК в этой клетке;
в) синтез клеткой-хозяином конечных продуктов на основе генетической информации рекомбинантной ДНК.
ЭВОЛЮЦИОННОЕ УЧЕНИЕ
ПРЕДПОСЫЛКИ ВОЗНИКНОВЕНИЯ ДАРВИНИЗМА
Объективные предпосылки
1. Завершение эпохи Великих географических открытий (многочисленные описания не известных ранее земель, их природных условий, флоры и фауны), разрушивших воззрения об однородности и неизменности мира.
2. Активная колонизация новых земель европейцами, которая требовала создания теории позволяющей определять целесообразность этих действий.
3. Бурное развитие капитализма требовало такого же от развития сырьевой базы, в том числе от сельского хозяйства, которое уже исчерпало метод «проб и ошибок».
4. Интенсивное развитие науки, в свою очередь, ускорило процесс создания эволюционного учения.
Трансформизм – система представлений естествоиспыта-телей и философов 17–19 веков об исторической изменяемос-ти (трансформации) организмов. Сторонники трансфоормиа-низма (Р.Гук, П.Мопертюи, Ж.Ламерти, Д.Дидро, Ж.Бюффон, И.В.Гёте, Эразм Дарвин, Э.Жоффруа Сент-Илер, К.Ф.Рулье, А.А.Каверзнев и др.) постулировали, но не доказывали эволюционные преобразования организмов.
Карл Линней в работах: «Система природы» и «Философия ботаники» (1735) дал определение вида: «Вид – это совокупность особей сходных как дети одних родителей или как родители и их дети». Он считал вид результатом отдельного акта Творца и отрицал существование эволюции (в конце жизни признал возможность изменения особей в результате скрещивания. К. Линней описал более 8000 видов, объединил сходные виды в роды, сходные роды – в отряды, а отряды – в классы и закрепил использование латинской бинарной номенклатуры. Систематику проводил по двум, трём наиболее ярким внешним признакам, поэтому допускал ошибки. К. Линней, сознавая искусственность своей системы писал: «Искусственная система служит только до тех пор, пока не найдена естественная».
Жан Батист Пьер Антуан де Монье шевалье де Ламарк в вводных лекциях по зоологии (1802 – 1806) и в труде «Философия зоологии» (1809) изложил теорию эволюции.
В подтверждение реальности существования эво-люции создал «Лестницу существ», на 6 ступенях (градациях) которой (по мере усложнения нервной и кровеносной систем) были расположены, выделенные им 14 классов животных. На 1-ой градации Ламарк поместил инфузорий и полипов, на 2-ой – лучистых и червей, на 3-ей – насекомых и паукообразных, на 4-ой – ракообразных, кольчатых червей, усоногих и моллюсков, на 5-ой – рыб и рептилий, на 6-ой – птиц, млекопитающих и человека.
Причиной эволюции Ж. Б. Ламарк считал изменение условий окружающей среды.
Движущую силу эволюции определял как внутреннее стремление организма к совершенству, к гармонии с природой и сформулировал два закона эволюции:
Закон упражнения и не упражнения: «Органы активно используемые организмом в течении жизни развиваются, а не используемые – деградируют и исчезают».
Закон безусловного наследования: «Благоприобретённые в течении жизни признаки обязательно передаются по наследству».
Карл Бэр в 1828 году сформулировал закон зародышевого сходства: «Зародыши всех позвоночных животных в процессе развития проходят сходные стадии развития, причём более высокоорганизованные организмы повторяют стадии менее организованных».
Чарльз Лайель в работе: «Основы геологии» (1830) говорил о непрерывно продолжающейся эволюции поверх-ности Земли, демонстрируя наличие прогрессивных изменений в ископаемых остатках.
Андрей Николаевич Бекетов в 1858 году установил взаимосвязь между вегетативным строением растений и средой их обитания.
Субъективные предпосылки
Эразм Дарвин - дед Чарльза Дарвина был одним из виднейших эволюционистов своего времени, он оказал боль-шое влияние на становление Ч. Дарвина, как учёного.
Кругосветное путешествие на корабле «Бигль» в ка-честве натуралиста (1831 –1836 гг.) позволило Ч. Дарвину самому увидеть разнообразные виды в естественных условиях их обитания.
Ч. Дарвин с детства обладал способностью и стремлением устанавливать причинно-следственные связи наблюдаемых им фактов и процессов.
Основные этапы создания теории эволюции
1839 г. Ч. Дарвин публикует отчёт о путешествии на корабле «Бигль».
1858 г. Ч. Дарвин получает рукопись статьи Альфреда Уоллеса, в которой тот формулирует основные принципы естественного отбора.
1858 г. Ч. Дарвин и А. Уоллес по настоятельной просьбе геолога Ч. Лайеля и ботаника Дж. Гукера делают доклады на заседании Линнеевского зоологического общества в Лондоне и вместе их публикуют в печати.
1859 г. Издаётся работа Ч. Дарвина «Происхождение ви-дов путём естественного отбора, или сохранение благоприятс-вуемых рас (форм, пород) в борьбе за жизнь».
1886 г. Ч. Дарвин публикует второй капитальный труд - «Изменение домашних животных и культурных растений».
1871 г. Выходит в свет третий труд, подводящий итог создания стройной теории эволюции: «Происхождение человека и половой отбор».