
М. В. Ломоносова биологический факультет на правах рукописи никольская кира Алексеевна системно-информационные аспекты познавательной деятельности позвоночных диссертация
| Вид материала | Диссертация |
| А – контрольная группа, отклонения относительно средней активности во всех структурах мозга; В |
- Московский государственный университет имени М. В. Ломоносова биологический факультет, 1266.34kb.
- М. В. Ломоносова биологический факультет н а правах рукописи Столяров Андрей Павлович, 657.12kb.
- Самовольцева Кира Алексеевна Дылевич Елена Валерьевна закон, 168.26kb.
- На правах рукописи Гуськова ирина Алексеевна, 682.76kb.
- На правах рукописи, 772.97kb.
- На правах рукописи, 315.69kb.
- На правах рукописи, 372.38kb.
- Тема: «Системно-деятельностный подход - универсальный инструмент познавательной деятельности», 64.45kb.
- Самовольцева Кира Алексеевна Агеева Елена Валерьевна закон, 118.95kb.
- На правах рукописи, 544.87kb.
Обнаруженная в фармакологических исследованиях четко выраженная разнонаправленность поведенческих эффектов под действием психостимулирующих препаратов у выделенных нами двух подгрупп III типа давала основание говорить о том, что физиологическое разнообразие (особенности основных нервных процессов), могло быть обусловлено биохимическими различиями. Достаточно демонстративными в этом отношении оказались психо-эмоциональные проявления (рис. 9А). При увеличении дозы психо-эмоциональная выраженность большинства реакций оказывалась инвертированной по сравнению с малыми дозами препарата и сохраняла свою несхожесть относительно двух подгрупп. Сходная зависимость направленности психо-эмоциональных эффектов у «сангвиников» и «флегматиков», обнаруженная при разрушении гиппокампа (см. раздел 6) указывали на то, что физиологические различия у этих двух групп животных обусловлены не только биохимическими, но и морфофункциональными различиями (рис. 9 В).
В целом, выявленные в исследовании такие особенности ПД крыс III типа, как низкая интенсивность ассоциативной активности, высокие интегративные возможности, способность оценивать ассоциативные связи не только по их семантической значимости (прогнозировать действия за счет S R связей), но и по их прагматической ценности (прогнозировать результат будущего действия за счет S1 S2 связей), важная роль перцептивного научения давали основание определить эту группу крыс как животных с концептуальным типом ПД. Наличие таких признаков, как отсутствие прямой зависимости результата интегративного процесса от предшествующего опыта, как это характерно для процедурного типа, высокая вероятность его проявления в поведении при минимальном числе апробирования, позволяли утверждать, что инсайтный (Келер, 1932), или когнитивный (по Торпу, 1963) тип обучения, вероятнее всего, присущ всем позвоночным, обладающих сходными с крысами III типа чертами познавательной деятельности.
Таким образом, на основе использования системно-информационного подхода при изучении индивидуальных особенностей ПД удалось выделить совокупность психофизиологических признаков, определяющих специфику трех познавательных типов: сигнального, процедурного и концептуального. Сигнальный тип способен был обучаться только по жесткому типу, устанавливая однозначное соответствие между раздражителем внешней среды и ответной реакцией организма - S→R связь. Процедурный тип способен был в условиях свободного выбора организовать поведение на основе формирования цепочек из элементарных связей. Концептуальный тип на основе компиляции из образованных цепочек действий мог формировать схему на основе Sk – Sn связей.
- биохимические корреляты типологии познавательной деятельности
Анализ состояния нейромедиаторных систем мозга выявил четкую структурно-обусловленную гетерогенность ответа этих систем на информационную нагрузку. Более сильная реакция со стороны сенсомоторной коры СМК по сравнению с хвостатым ядром ХЯ свидетельствовали о глубокой вовлеченности коры в реализацию результата ПД. При этом выяснилось, что направленность сдвигов активности медиаторов и ферментов их утилизации не зависела от особенностей ПД и носила исключительно количественный характер: чем выше был уровень ПД, тем большие сдвиги наблюдались в активности медиаторных систем.
Резкое увеличение содержания норадреналина НА (в 3 раза) в сенсомоторной коре при низкой чувствительности дофамина ДА и МАО Б к действию информационной нагрузки, свидетельствовали об активации катехоламинергической КА-системы. Сходная тенденция была выявлена и для холинергической АХ-системы: увеличение активности фермента синтеза ацетилхолина – ацетилхолинтрансфераза ХАТ при значимом падении активности ацетилхолинэстеразы АХЭ приводило к выраженному возрастанию содержания свободного АХ в коре. Активация холинергической системы в хвостатом ядре согласно обменному коэффициенту – ХАТ/АХЭ проявилась в повышении оборота этого медиатора. Степень сдвигов в этих двух нейромедиаторных системах коррелировала с уровнем двигательной активности. Реакция со стороны серотонинергической системы (5-ОТ) была обнаружена только в СМК и состояла в сходном увеличении содержания серотонина у животных, кардинально отличавшихся по уровню познавательной деятельности. В отличие от КА- и АХ- систем, наибольшие изменения в 5-ОТ-системе были характерны для животных с низким уровнем ПД. Наличие сходного ответа нейромедиаторных систем у крыс с разной степенью влечения к алкоголю позволяет говорить о том, что нейрохимические сдвиги отражают общий принцип реагирования на фактор вовлеченности ЦНС в когнитивную деятельность, демонстрируя многократно описанную реципрокность функциональных взаимоотношений 5-ОТ-системы с КА- и АХ- системами (Семенова, 1976; Громова, 1980; Кругликов, 1986; Pujol et al., 1973; Reaux-Le Goazigo, et al., 2005; de Mota et al., 2008).
Исследование активности ферментов белкового и углеводного обмена при обучении выявило устойчивые изменения их активности, направленность которых зависела от чувствительности ферментов к действию информационной нагрузки и индивидуальных особенностей ПД животных (рис. 10).
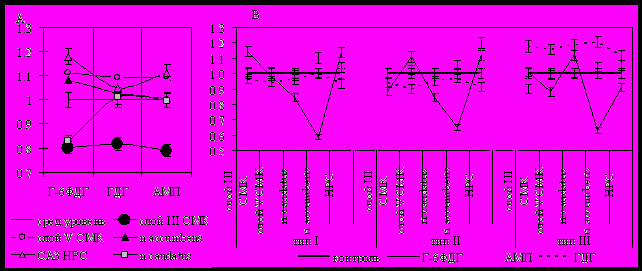
Рис. 10. Активность ферментов углеводного и белкового обмена в структурах мозга у крыс Вистар, различающихся по характеру познавательной деятельности.
А – контрольная группа, отклонения относительно средней активности во всех структурах мозга; В – группы животных с различным типом ПД: I – сигнальный тип, II –процедурный тип; III – концептуальный тип; по вертикали – отклонения активности ферментов в каждой структуре мозга по отношению к исходному уровню в отн. ед.
При наличии исходно высокой межструктурной вариабельности активности ферментов у интактных животных (рис. 9А) удалось установить, что наибольшая чувствительность у всех животных, независимо от их типологических особенностей ПД, была характерна для глюкозо-6-фосфатдегидрогеназы Г-6ФДГ – ключевого фермента пентозофосфатного шунта, а наименьшая – для аминопептидазы АМП, отщепляющей N-концевые аминокислотные остатки от белков и пептидов и способной гидролизовать различные дипептиды. В отличие от первых двух ферментов реакция со стороны глутаматдегидрогеназы ГДГ, играющей важную роль в энергетическом обмене веществ, была связана исключительно с характером решения проблемной ситуации, направленность которой зависела от особенностей ПД (рис. 10 В). В отличие от сигнального типа (I), реакция ГДГ в виде снижения активности у процедурного типа (II) наблюдалось только в эфферентных нейронах СМК и гиппокампе. В тоже время для концептуального типа (III) резкое смещение активности ГДГ в область высоких значений во всех изученных нами структурах, свидетельствовало о широкой структурной заинтересованности в более высоком энергетическом обеспечении при обучении (рис. 10 В).
Однонаправленность реакции Г-6ФДГ в виде резкого снижения его активности в n. аccumbens в ответ на процедуру обучения у животных, различавшихся не только по уровню двигательной, но и эмоциональной и когнитивной активностям, позволяет рассматривать реакцию этой структуры как маркер, отражающий факт вовлечения ЦНС в познавательный процесс. Полученные факты подтверждают представление о прилежащем ядре как ключевой структуре в интеграции функций лимбических и стриарных отделов мозга при запуске как врожденных двигательных форм поведения, так и более общих процессов, таких как внимание (Громова, 1980; Данилова, 1992; Yechenko, 2004). Характер и степень различий (73.3%) в структурном распределении активности изученных ферментов, обнаруженных у двух обучившихся групп, позволяют говорить о том, что рисунок вовлеченных структур мозга отражает не столько уровень двигательной активности (Герштейн, 1971; Сергутина, 1989), поскольку по этому признаку они были сходны на этапе, предшествующем цитохимическому анализу, сколько специфику структурно-химической топографии энергетической вовлеченности ЦНС в обеспечение реализации познавательной активности.
Изучение состояния белкового обмена в нейронах СМК, дорсального НРС, ХЯ и амигдале (n. ABL) показало, что направленность изменений и уровень отклонений были тесно связаны с особенностями ПД животного. У процедурного типа (II тип) изменения в СМК были выражены крайне слабо и зависели от функциональной принадлежности нейронов: в афферентных нейронах снижение содержания белка наблюдалось только в ядрах клеток, а в эфферентных нейронах реакция оказывалась более системной, поскольку содержание белка увеличивалось как в ядрах, так и в цитоплазме клеток (рис. 11). Реакция со стороны лимбических структур на вовлеченность животных в познавательный процесс, в отличие от СМК, оказалась весьма специфичной. Под действием информационной нагрузки исходно существующие цитохимические различия в нейронах лимбических структур стирались и их функциональное состояние приобретало однотипный ответ. Резкое однонаправленное снижение всех морфо-химических показателей состояния белкового обмена указывает на то, что не только СМК, но гиппокамп и миндалина активно вовлечены в общий процесс, связанный с реализацией результата ПД (рис. 11), подтверждая правомочность представления о тесном взаимодействии «информационных» (кора – гиппокамп) и «мотивационных» (миндалина – гипоталамус) систем мозга при реализации поведения (Симонов, 2004; Swanson, 1988; Le Doux, 1989; Machado и J. Bachevalier, 2006).
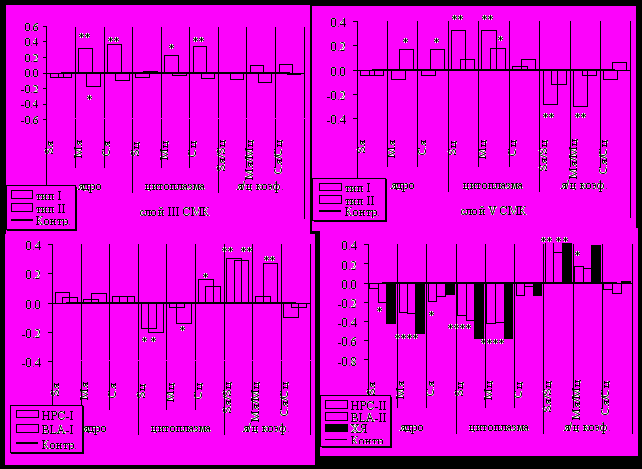
Рис. 11. Особенности состояния белкового обмена в структурах мозга у крыс Вистар, вызванные процессом обучения.
СМК – сенсомоторная кора, НРС – дорсальный гиппокамп, ABL – базолатеральное ядро амигдалы, ХЯ – хвостатое ядро; по вертикали – значения площади (S), содержания (М) и концентрации (С) белка в ядре (я) и цитоплазме (ц) нейронов, а также ядерно-цитоплазменных коэффициентов относительно контроля, принятого за ноль.
Характер морфохимических сдвигов у крыс с подавленными признаками ПД (сигнальный, I тип) имел противоположную направленность в изученных нами структурах. При этом высокий уровень пассивных стрессовых реакций, повышенная склонность животных к формированию реакции предпочтения к этанолу, указывали на наличие устойчивого стрессового состояния, снижавшегося при потреблении раствора этанола. Негативная реакция на когнитивную нагрузку в афферентных нейронах СМК выразилась в резком возрастании содержания и концентрации белка в ядре и цитоплазме при сохранности их размеров, при этом реакция со стороны ядра на фоне выраженной дегидратации была достоверно выше. В эфферентных нейронах СМК (слой V) значимое повешение размеров и содержания белка было установлено лишь для цитоплазмы клеток (рис. 10). Важным, на наш взгляд, является тот факт, что ответ нейронов НРС и n. ABL на когнитивную нагрузку у этих животных, несмотря на негативный результат обучения, был сходен с таковым у крыс, успешно решивших задачу.
Изучение состояния белкового обмена в нейронах хвостатого ядра показало, что эта структура активно вовлечена в реализацию ПД (рис. 11). В ассоциативных нейронах это проявилась в резком снижении всех морфохимических показателей как в ядре, так и в цитоплазме. Сопоставление реакции ХЯ с таковой в лимбических структурах, несмотря на отличия в двигательной, когнитивной и психо-эмоциональной сферах, выявило существенное сходство. Однонаправленность изменений, обнаруживаемая не только в отношении состояния белкового обмена в нейронах ХЯ, НРС и n. BLA, но и в структурной топографии ферментативной активности (отличие только по 3 из 15 показателей, рис. 10 В) и нейромедиаторных реакциях свидетельствуют о том, что эти изменения обусловлены единым психофизиологическим процессом. В рамках концепции о ведущей роли стриопаллидарной и лимбической систем мозга в механизмах внимания, (Суворов, 1980; Данилова, 1992 Grossberg, 1982; Grossberg, 1990;

1 - ИИА – скорость нарастания ошибок, VУГ – скорость снижения числа ошибок с максимально достигнутого до устойчивого минимума, СИН – степень инертности = кол-во нерациональных реализаций УР, потребовавшихся для перехода к минимизированному выполнению; РПЭ – доля психо-эмоциональных реакций в общем объеме, совершенных за 15 опытов; Д –длительность. И –интенсивность, С – сила, УРВ и УРТ – условнорефлекторное возбуждение и торможение. Данные приведены относительно контроля, принятого за единицу, значения ЗК в контроле приняты за ноль; р = * - 0.05 и ** - 0.001 – достоверные значения по сравнению с контролем в соответствие с критерием t-Student.
Такие факты, как отсутствие активных психо-эмоциональных проявлений при совершении ошибки в условиях высокой их плотности в пределах опыта, способность обучаться в условиях исключительно низких, вдвое ниже контроля, – M ± m = 0.16 ± 0.01 значений вероятности успеха на фоне эмоциональной уплощенности свидетельствовали о снижении чувствительности к факту неподкрепления. Сделанное заключение согласуется с представлениями (Прибрам, 1972; Пигарева, 1974; Симонов, 2000), согласно которым в результате гиппокампэктомии могут изменяться критерии оценки происходящих событий.
Повреждение НРС у крыс с концептуальным типом ПД, в отличие от первых двух, сопровождалось эффектами, знак и направленность которых зависели от исходных возбудительно-тормозных соотношений. В случае умеренного преобладания возбуждения - IIIА тип (сангвинические черты) операция повлияла, в основном, на возбудительный процесс, сократив продолжительность, но при этом, резко усилив его интенсивность – число выполнений в единицу времени. Эти изменения в сочетании с сокращением продолжительности тормозного процесса негативно сказались на характере проявления внутреннего торможения (ДУРТ, ИУРТ и СУРТ, табл. 4), свидетельствуя о резком снижении функции внимания. Наиболее выраженные последствия операции у животных этого типа выявлялись на этапе реализации сформированного навыка. Высокая неустойчивость УР поведения за счет резкого снижения длительности и интенсивности УР торможения сочеталась с усилившейся инертностью нервных процессов (табл. 4).
В случае преобладания тормозного процесса (IIIИ тип, флегматические черты), операция привела к значимому усилению возбудительного процесса, провоцируя смещение соотношения нервных процессов в сторону возбуждения. Это обусловило снижение пассивных проявлений, особенно на начальных этапах обучения, за счет чего процесс обучения, в отличие от контроля, сопровождался психо-эмоциональными реакциями преимущественно раздражительного характера. Проведенные исследования показали, что основные изменения у крыс этого типа при сохранности всех специфических признаков осуществления ПД были связаны с усилением инерционных свойств ЦНС. Навязчивое повторение одних и тех же ошибочных действий, прежде чем животные переходили к выполнению других вариантов, давали основание рассматривать эти факты как доказательство снижения функциональной лабильности очагов возбуждения, обеспечивающих реализацию УР процесса. Наиболее четко эти признаки выявлялись на этапе стабилизации навыка и в пробах с переделкой навыка. Обнаруженное усиление инерционных свойств ЦНС можно рассматривать как доказательство сужения объема внимания, с одной стороны, и снижение скорости извлечения зафиксированного события из долговременной памяти (Крюков, 2002), с другой стороны, что многими исследователями в случае оценки памяти по количеству затраченных проб/выполнений или опытов может восприниматься как ее серьезное ухудшение.
Большой интерес к амигдале в последнее время вызван данными о ее связях с различными гиппокампальными областями (Amaral et al., 1992; Hamann et al., 1999; Pitkänen et al., 2000), на основании чего высказано представление о важной роли амигдало-гиппокампальных связей в обучении и памяти (Bonda et al., 1996; Zald et al., 1997; Cahill et al., 1998, Armony,

1 - ИИА – скорость нарастания ошибок, VУГ – скорость снижения числа ошибок с максимально достигнутого до устойчивого минимума, РЭ – доля психо-эмоциональных реакций в общем объеме, совершенных за 15 опытов; все данные приведены относительно контроля, принятого за единицу; р = * - 0.05 и ** - 0.001 – достоверные значения в соответствии с критерием t-Student.
Результаты исследования показали, что операция у всех крыс, как и в случае дорсального НРС, серьезно повлияла на характер проявления основных нервных процессов. Гипервозбудимость – специфический признак амигдалэктомированных животных, обусловил двигательную гиперактивность, наиболее ярко проявлявшуюся в ситуации новизны. При этом амигдалэктомия, в отличие от НРС, спровоцировала увеличение не только уровня возбуждения и глубины торможения, но и скорости выхода из тормозного состояния, обеспечивая, тем самым, «импульсивный» характер развития возбудительно-тормозных процессов. Неуравновешенность нервных процессов в первую очередь отразилась на характере психо-эмоциональных проявлений животных (табл. 5). В поведении животных, независимо от индивидуальных особенностей, доминировали реакции раздражительного типа преимущественно в виде различных смещенных и стрессовых проявлений. При этом, в отличие от повреждения НРС, операция не сопровождалась явлениями эмоциональной уплощенности и отсутствием эмоционального резонанса.
Спровоцированные амигдалэктомией сдвиги в возбудительно-тормозной сфере негативно сказались как на скорости осуществления, так и характере реализации ПД, направленность которых зависела от индивидуальных особенностей животного (табл. 5). Способность амигдалэктомированных крыс (АМ-крыс) сигнального типа уже в 4.47 ± 0.06 опытах (p < 0.01) сформировать ассоциацию, соответствующую структуре задачи при наличии сниженной тревожности свидетельствовала об облегчающем влиянии операции на процесс восприятия за счет интенсификации ассоциативных свойств. Минимальное количество проб (18.6 ± 0.56) оказалось достаточным, чтобы в 62.4 % случаев воспроизвести ее с первой пробы в следующем опыте (через 24 ч). Устойчиво генерализованный характер посещения всех семантических зон лабиринта в дальнейшем не смог завершится образованием навыка.
Наиболее серьезно амигдалэктомия оказала влияние на крыс с процедурным типом ПД. Усиление возбудительного процесса у исходно возбудимых животных, двигательная гиперактивность, возникавшая в ситуации новизны и в начале каждого опыта, а также при когнитивном напряжении, импульсивный характер развития обоих нервных процессов указывали на появление дефектов в работе механизмов, обеспечивающих устойчивость активационной системы ЦНС. Повышенная при этом скорость перехода с одного нервного процесса на другой придавали поведению генерализованный характер. Незавершенность действий, низкая вероятность подкрепления, многократные повторы одних и тех же действий отразились на значениях когнитивных затрат (табл. 5). Многократные, а подчас навязчивые выполнения одних и тех же последовательностей действий, часто ошибочных, способствовали запечатлению этих ассоциаций, которые в дальнейшем постоянно воспроизводились. Несмотря на крайне высокий уровень «шума» (Русп = 0.11 ± 0.02), АМ-крысы смогли распознать значимые элементы задачи и образовать корректную ассоциацию, соответствующую решению задачи. Такие факты, как: (1) наличие инстинктивной программы – ВхК1+Вых1… в начале каждого выполнения пищедобывательной задачи (Р = 0.83 ± 0.02 вместо 0.14 ± 0.01 в контроле; р < 0.01), (2) когнитивная генерализация, (3) возможность воспроизведения адекватного решения - ВхК1К2Вых только на пике эмоционального напряжения указывали на серьезные трудности, связанные с концентрацией УР возбуждения, обусловленные ослаблением внутреннего торможения (табл. 5).
Эффекты амигдалэктомии у крыс с концептуальным типом ПД (IIIА и IIIи типы) оказались не столь драматичными как у процедурного типа, степень выраженности и направленность которых, как и у НРС-крыс этого типа, во многом зависела от исходного соотношения возбудительно-тормозных процессов. В случае незначительного превалирования возбуждения (подгруппа IIIА), амигдалэктомия в поведенческой и эмоциональной сферах провоцировала сходные с процедурным типом изменения за счет резкого усиления возбудительного процесса, хотя уровень двигательной гиперактивности был намного слабее. Поскольку операция не затрагивала тормозные характеристики, угашение ошибочных и лишних действий развивалось в соответствие с контролем. Когнитивный процесс у АМ-крыс этого типа практически не был затронут (табл. 5). Существенное ослабление всех параметров УР торможения на фоне явного усиления возбудительного процесса негативно сказалось на характеристиках воспроизведения памятного следа. Процесс организации УР очага (концентрация возбуждения), необходимый для активизации памятного следа и подключения следящей системы (внимания) у крыс IIIА типа осуществлялся с большим трудом после операции.
У животных флегматического типа (IIIИ) амигдалэктомия привела к более серьезным психофизиологическим последствиям (табл. 5). При сходстве большинства изменений в области проявления основных нервных процессов, эмоциональных реакций, когнитивного процесса и поведенческих характеристик, организация навыка была существенно затруднена. Значимое сокращение продолжительности обоих нервных процессов на фоне резких переходов с одного процесса на другой (специфическая комбинация изменений, характерная только для этого типа) негативно сказалось не только на проявлении УР торможения, но и на показателях УР возбуждения (табл. 5). Несмотря на интенсификацию ассоциативного и интегративного процессов, спровоцированную амигдалэктомией, присутствие в навыке большого числа примитивных тактик, низкая организованность навыка – все эти факты, обнаруженные у АМ-крыс концептуального типа, свидетельствовали о серьезном нарушении механизмов, обеспечивающих организацию и устойчивость воспроизведения памятного следа
Таким образом, благодаря использованию системно-информационного подхода удалось получить убедительные свидетельства того, что дорсальный НРС и амигдала не имеют прямого отношения к реализации когнитивных процессов и функционированию памяти. Несоответствие полученных результатов данным литературы, по нашему мнению, обусловлено отсутствием в большинстве работ системного подхода при изучении поведенческих эффектов и системы оценок, позволяющих раскрыть содержательную сторону поведения - что и как делало животное. В целом, выявленная нами ведущая роль возбудительно-тормозных отношений в проявлении индивидуальных черт, значимый вклад в которые оказывали и гиппокамп и амигдала, свидетельствовала о том, что эти две структуры входят в единую активирующую систему ЦНС, ответственную за оперативную организацию доминанты и условия ее функционирования (Ухтомский, 1966; Крюков, 2002), обеспечивая тем самым осуществление акта внимания. На основании полученных результатов высказано представление о том, что благодаря этой активирующей системы обеспечивается: (1) ее устойчивость за счет сдерживающих амигдалярных влияний от чрезмерного и неуправляемого распространения возбуждения из фокуса доминанты; и (2) функциональная лабильность за счет возбуждающих гиппокампальных влияний, позволяющих в пределах доминантной системы переключаться с одного локального центра на другой. Сделанное нами заключение возвращает к представлениям о важной роли гиппокампа и амигдалы в осуществлении процессов возбуждения и торможения, популярных в 60-80 г.г. прошлого столетия (Меринг, 1974; Carmos, Grastyan, 1962; Kimble et al, 1966), к которым исследователи последних лет стали все чаще возвращаться при объяснении эффектов гиппокампэктомии и амигдалэктомии (Jarrard, et al, 1998; Sainsbury, 1998; Chan, et al, 2001).
- Эволюционные аспекты познавательной деятельности позвоночных
Отсутствие четких представлений об эволюции ПД животных подтолкнула нас использовать системно-информационный подход для изучения этого вопроса на примере позвоночных, обладающих разным уровнем организации мозга и обитающих в различных экологических условиях.
Проведенные исследования выявили, что в каждой из изученных групп позвоночных за исключением вторичноводных имелись особи, которые смогли в условиях свободного выбора решить предложенную проблемную ситуацию и сформировать устойчивый пищедобывательный навык. Процент успешного решения задачи в пределах каждой группы позвоночных был видоспецифичен (рис.13).
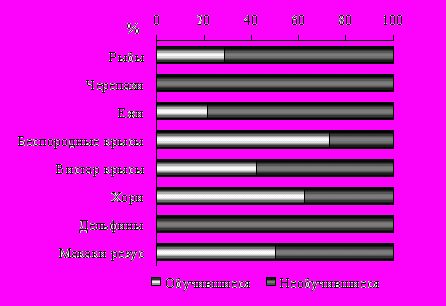
Рис. 13. Соотношение животных, решивших и отказавшихся от решения пищедобывательной задачи в ряду позвоночных.
Изучение ПД наземных млекопитающих показало, что оценка процесса обучения с помощью традиционных показателей – проб и опытов не выявила серьезных различий в скорости обучения в ряду насекомоядные → обезьяны, подтвердив правомерность представлений о независимости ПД от морфо-функциональной организации мозга (Макфайл, 1986, 2001). С помощью системно-информационного анализа удалось показать, что процесс распознавания синтаксической и семантической информации, независимо от уровня организации мозга, начинался с образования одинакового конечного объема «алфавита» в виде двуграмм, формирование которого заканчивалось у всех видов ко 2-3-му опыту (табл. 1). Однако, имея изначально сходный объем «двигательного алфавита», интенсивность ассоциативного процесса была тем выше, чем лучше были развиты ассоциативные системы мозга в ряду наземных млекопитающих, тем выше была скорость распознавания структуры задачи, тем выше был уровень представления о структуре пространства. Чем выше был уровень цефализации, тем легче животное улавливало признаки симметрии в лабиринте, тем чаще оно прибегало к операции зеркального переноса информации при моделировании «карты» всего пространства. Высокая скорость ассоциирования у высших млекопитающих, скачкообразное образование длинных маршрутов по всему лабиринту, включая побежки между подкрепляемыми кормушками уже во втором опыте, активное использование операции переноса свидетельствовали о более высокой интенсивности ассоциативного процесса (табл. 1).
Особенности становления целенаправленного поведения проявились в том, что: (1) чем выше был уровень организации мозга, тем продолжительнее был этап хаотического поведения (большая доля таких проб в общем объеме), тем меньшую роль играли внешние ориентиры для реализации цели и (2) тем быстрее ориентация в пространстве осуществлялась на основе векторного представления о пространстве. При этом, если у насекомоядных и грызунов переход от одного этапа обучения к другому был четко выражен, то трансформация стохастического поведения в целенаправленное у хищников и обезьян приобретала скачкообразный характер, за счет чего феномен целенаправленности проявлялся по логистическому типу.
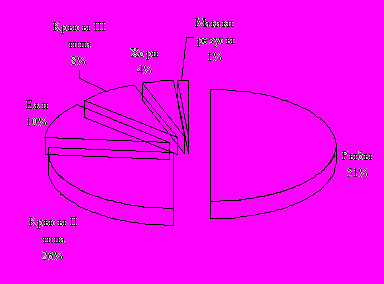
Рис. 14. Когнитивные затраты (КЗ), потребовавшиеся наземным млекопитающим на распознавание условий задачи и формирование навыка. Значения КЗ для каждой группы концептуального типа показаны в виде доли относительно общей суммы КЗ (принятой за 100%).
С интенсификацией ассоциативного процесса в ряду наземных млекопитающих улучшались и интегративные показатели. Выяснилось, что, чем шире были представлены ассоциативные системы в ЦНС млекопитающего, тем меньше нужно было животному совершить повторных попыток при одном и том же объеме подкрепления, чтобы распознать условие задачи. Хотя тестируемые группы мало различались по числу затраченных проб (заходов в лабиринт), переход на когнитивно-обусловленные формы ответа происходил существенно быстрее, чем выше на эволюционной лестнице находилось животное. Эта тенденция четко проявилась в показателе познавательных "затрат", потребовавшихся на формирование целенаправленного поведения (рис. 14). По мере усложнения организации переднего мозга все четче проявлялся феномен абстрагирования, когда роль подкрепления начинали выполнять не только физические ощущения от контакта с пищей, но и психические (сенсорная фиксация наличия пищи в кормушке). Увеличение числа таких проб в ряду наземных млекопитающих, особенно у обезьян, свидетельствовало о возрастающей роли психического фактора.
Исследование ПД первичноводных позвоночных (Cyprinidae: Carassius auratus и Cyprinus carpio) показало, что рыбы при использовании типичных для наземных млекопитающих методических условий: ТОП = 20с и межопытного интервала МОИ = 24 ч не способны решить предложенную задачу в условиях свободного выбора. Используя процедуру последовательного увеличения ТОП и МОИ удалось показать, что аналитико-синтетический процесс у рыб, направленный на решение задачи и формирование целенаправленного навыка смог осуществиться только при достижении 60-мин ТОП при 48-часовом МОИ. Характерно, что обучение (по числу проб), как и у млекопитающих, развивалось по логистическому типу. Начиная с 12-го опыта, скачкообразно возрастала “производительность работы” в опыте, рыбы стереотипно могли осуществлять навык с частотой 2.26 ± 0.15 пробы в мин без каких-либо признаков усталости и стрессируемости. Возможность осуществления рыбами ПД, сходной с млекопитающими, только при очень длительной экспозиции информации, свидетельствовали о низкой скорости информационного процессинга, обусловленной, вероятнее всего, низкими «энергетическими» возможностями ЦНС.
Несмотря на единую с млекопитающими динамику распознавания различных видов информации, характер организации программы поведения у рыб свидетельствовал о серьезных отличиях от такового у наземных млекопитающих. Выяснилось, что ведущую роль у рыб в построении навыка играла инстинктивная программа (ВхКВых). Выполненная на одной половине лабиринта, она зеркально воспроизводилась на другой для посещения второй кормушки в пределах одной пробы, хотя существовал прямой переход между кормушками (см. рис. 1, Л3, вариант с заштрихованными областями). Такие факты как: (1) независимость формируемой программы навыка от знания пространственной и семантической информации; (2) включение внешних ориентиров в качестве направляющих сигналов в структуру навыка; (3) ситуационно-обусловленный обход как завершающий этап обучения, а не начальный как у наземных млекопитающих – позволяет предположить, что когнитивный процесс и организация навыка для рыб являются двумя независимыми задачами. Тот факт, что на фоне стереотипного выполнения навыка самопроизвольно с вероятностью M ± m = 0.42 ± 0.06 стал оперативно воспроизводиться сходный с млекопитающими когнитивно обусловленный вариант решения задачи - Вх К1 К2 Вых позволяет говорить о том, что рыбы способны к обобщению. В целом, полученные факты свидетельствуют о том, что, проявляя оперативно высокие познавательные способности, рыбы не могли использовать их (как опыт) для последующей организации адекватного поведения. Обнаруженные особенности решения предложенной когнитивной задачи обуславливали высокие значения познавательных "затрат" у рыб (рис. 14).
Отсутствие четких представлений о причинах, побудивших древних представителей наземных позвоночных вернуться в водную среду, давало основание исследовать особенности ПД на примере двух видов вторичноводных позвоночных, типичных объектах экспериментальной психологии – болотных черепахах (Clemmys caspica) и дельфинах (Tursiops truncatus).
Результаты обучения черепах в условиях свободного выбора показали, что они, в отличие от рыб, не смогли сформировать целенаправленное поведение ни в сходной с рыбами лабиринтной ситуации при длительности опыта от 20 до 60 мин, ни в более простой задаче на чередование в среде К2 (см. раздел Методика). Столкнувшись с тем, что в ситуации чередования животные с первой попытки осуществляли прямой переход между кормушками, мы модифицировали условия задачи таким образом, чтобы подкреплялся любой обходной вариант кроме прямого, который был перекрыт. Черепахи, в отличие от грызунов, в измененных условиях не справились с такой задачей и смогли обучиться только по методике жесткого обусловливания. При повторном открытии перегородок после закрепления нового маршрута (234.5 ± 28.7 проб) животные сразу же возвратились к использованию кратчайшего маршрута, игнорируя факт неподкрепления. Полученные факты: (1) полное отсутствие исследовательской активности при попадании в незнакомую среду, независимо от степени ее топологической сложности; (2) быстрое и практически полное подавление двигательной активности после выполнения первых негативных проб; (3) неспособность затормозить инстинктивную программу – указывали на серьезные дефекты у черепах в сфере прогностических способностей.
Специфика работы с дельфинами также потребовала видоизменить структуру экспериментальной среды: при сохранности многоальтернативности лабиринтной среды, задача в когнитивном отношении была упрощена, поскольку имела только одно правильное решение. Предварительно дельфины были обучены элементарным навыкам – проходу через калитку, воздействию рострумом на различные предметы. Дельфинов тестировали в двух экспериментальных ситуациях: в лабиринтной среде (эксперимент 1) животные должны были сформировать 6-звенный пищедобывательный навык в циклической форме, в эксперименте 2 – 4-звенный навык с выходом в открытую морскую акваторию.
Проведенные исследования показали, что все животные в условиях свободного выбора, как и черепахи, отказывались решать пищедобывательную задачу, хотя и обладали всеми необходимыми предварительно сформированными элементарными оперантными навыками. Такие факты, как: (1) жесткая привязка двигательного опыта к конкретной обстановке (отсутствие феномена переноса), (2) неспособность перестраивать сформированный навык при неожиданном изменении условий эксперимента, (3) неспособность дельфинов самопроизвольно использовать эхолокацию для поиска цели; (4) использование ее только в ответ на внешний звуковой стимул, (4) развитие в условиях свободного выбора устойчивого невротического состояния; но в тоже время (5) возможность быстрого формирования многозвенного навыка (до 6 звеньев в течение 2 опытов) в условиях жесткого обусловливания – все это указывало на то, что дельфины испытывают серьезные трудности при необходимости прогнозировать действие (ситуация выбора) и тем более прогнозировать результат действия.
Таким образом, экспериментальный материал, полученный на примере изучения ПД у первичноводных и наземных млекопитающих, позволяет высказать представление о том, эволюция ПД шла по пути увеличения сложности операционных программ (селекции и компиляции). Фактор преодоления проблемной ситуации решался через разнообразное сенсомоторное взаимодействие с внешней средой. Обучение в стационарных условиях опыта показало сходную динамику у всех этих животных, однако эффективность ПД зависела от сложности сенсорных, ассоциативных и исполнительных нервных систем, и, что наиболее важно, от уровня согласованности между ними. Чем больше была степень кортикализации этих систем, тем выше оказывалась скорость обработки информации и меньше когнитивная "стоимость" для того, чтобы решить задачу и сформировать правильную поведенческую программу, и тем больше был вклад перцептивного обучения в общий процесс обучения. Отличительные черты ПД изученных вторичноводных, позволяет говорить о том, что, несмотря на существенные различия в уровне организации их мозга, процесс познавательной эволюции этих животных преимущественно шел по пути усложнения ассоциативных механизмов.