
Научно-образовательный комплекс по кредитной технологии обучения опорные конспекты лекций по дисциплине "Биохимия" для студентов 1 курса специальности 050701 «Биотехнология»
| Вид материала | Конспект |
- Научно-образовательный комплекс специальности 050701 «Биотехнология» опорные конспекты, 386.71kb.
- Научно-образовательный комплекс по кредитной технологии обучения опорные конспекты, 429.71kb.
- Научно-образовательный комплекс По специальности 050701 «Биотехнология» учебно-методический, 644.78kb.
- Опорные конспекты лекций по дисциплине «биохимия» для студентов специальности 050727, 866.54kb.
- Научно-образовательный комплекс по специальности 050701 «Биотехнология» учебно-методический, 837.91kb.
- Научно-образовательный комплекс по специальности 050701 «Биотехнология» Методические, 834.35kb.
- Директор Инженерной Академии Док вет наук, проф. Е. Б. Никитин 2009 г. Автор: преподаватель, 828.3kb.
- Научно-образовательный комплекс по специальности 6N0701 «Биотехнология» учебно-методический, 704kb.
- Директор Инженерной Академии Док вет наук, проф. Е. Б. Никитин 2009 г. Автор: преподаватель, 952.2kb.
- Конспект лекций по дисциплине «сетевые технологии» (дополненная версия) для студентов, 2520.9kb.

МИНИСТЕРСТВО ОБРАЗОВАНИЯ И НАУКИ
РЕСПУБЛИКИ КАЗАХСТАН
ИННОВАЦИОННЫЙ ЕВРАЗИЙСКИЙ УНИВЕРСИТЕТ
Научно-образовательный комплекс
по кредитной технологии обучения
ОПОРНЫЕ КОНСПЕКТЫ ЛЕКЦИЙ
по дисциплине “Биохимия“
для студентов 1 курса
специальности 050701 «Биотехнология»
ПАВЛОДАР 2009
УТВЕРЖДЕНО
Директор Инженерной Академии
Док. вет. наук, проф. _________ Е.Б. Никитин
«___» _______________ 2009 г.
| Составитель: преподаватель | ______________Т.Н. Дубровина |
Кафедра “ Прикладная биотехнология”
Опорные конспекты лекций
по дисциплине
“Биохимия“
для студентов специальности 050701 «Биотехнология»
для очной формы обучения
на базе общего среднего образования
Опорные конспекты лекций разработаны на основании Государственного общеобязательного стандарта высшего образования РК специальности 050701 – «Биотехнология», ГОСО РК 3.08.327-2006, «Образование высшее профессиональное. Бакалавриат», г. Астана, 2006 г. , типовой учебной программы по дисциплине «Биохимия», утвержденной приказом МОН РК от 23.12.05 г. № 779 и рабочей программы 2008 г.
Опорные конспекты лекций рассмотрены на заседании кафедры «Прикладная биотехнология»
Протокол № ____ от _________200 г.
Зам. зав. кафедрой «Прикладная биотехнология» ________ М.С. Омаров
Опорные конспекты лекций утверждены на заседании научно-методического совета Инженерной Академии и рекомендованы к изданию
Протокол № ____ от __________200 г.
Председатель НМС Инженерной Академии
Канд. техн. наук, проф. ______________ Е.К. Ордабаев
Согласовано:
Начальник ИМО
к.п.н., проф. ________________ Н.М. Ушакова
сдано в медиатеку __________________
ТЕМАТИЧЕСКИЙ ПЛАН ЛЕКЦИЙ
| № | Темы лекций | Кол-во часов |
| Модуль 1 | ||
| 1 | Введение. Формирование структуры белка | 2 |
| 2 | Состав, структура и свойства ДНК и РНК | 2 |
| 3 | Структура и кинетические характеристики ферментов | 2 |
| 4 | Витамины | 2 |
| Модуль 2 | ||
| 5 | Обмен веществ | 2 |
| 6 | Углеводы | 2 |
| 7 | Липиды | 3 |
| Итого: | 15 | |
- Введение. Формирование структуры белка
Биохимия - это наука о качественном составе, количественном содержании и преобразованиях в жизненных процессах соединений образующих живую материю.
Предметом исследования биохимии являются метаболические процессы в клетке, а также регуляторные механизмы на уровне целостного организма. Метаболический процесс – это совокупность ферментативных реакций, катализирующих последовательность превращений реагенного соединения. Рассматрим процессы протекающие в живых организмах:
- Обмен веществ между организмом и внешней средой является обязательным условием существования живого организма.
- Живой организм обладает способностью реагировать на внешние воздействия веществ и таким образом приспосабливаться к изменениям во внешней среде.
- Для живого организма присуще способность к размножению и передаче по наследству характерных признаков строения и обмена веществ.
- Качественно новой ступенью в развитии материи является строгая упорядоченность протекания биохимических процессов, связанная с особенностями биологических структур.
Биохимия делится на несколько разделов: статическая, динамическая и функциональная.
Статическая - изучает химический состав живых организмов и свойства химических соединений. Функциональная - изучает биохимические процессы, лежащие в основе жизнедеятельности отдельных тканей и органов. Динамическая – изучает многообразные химические реакции, их взаимосвязь и регуляцию.
Белки
Белками, или белковыми веществами (протеинами, от греч. protas — первый, важнейший), называют высокомолекулярные (молекулярная масса варьирует от 5—10 тыс. до 1 млн. и более) природные полимеры, молекулы которых построены из остатков аминокислот. Каждый белок обладает своей, присущей ему последовательностью расположения аминокислотных остатков.
Биологические функции белков крайне разнообразны. Они выполняют каталитические (ферменты), регуляторные (гормоны), структурные (коллаген, фиброин), двигательные (миозин), транспортные (гемоглобин, миоглобин), защитные (иммуноглобулины, интерферон), запасные (казеин, альбумин, глиадин, зеин) и другие функции. Среди белков встречаются антибиотики и вещества, оказывающие токсическое действие.
Строение и аминокислотный состав белков
Аминокислоты — это гетеро-функциональные соединения. В молекуле аминокислоты содержится несколько функциональных (определяющих их свойства) групп: аминогруппа —NH2, карбоксильная группа —СООН и радикалы (остатки) —R, имеющие различное строение:
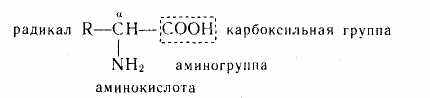
Число функциональных групп может быть различным: моно-аминомонокарбоновые, диаминомонокарбоновые, моноаминодикарбоновые аминокислоты и т. д. По строению боковых цепей (полярности, заряженности) аминокислоты группируются на неполярные (гидрофобные), полярные (гидрофильные), но не заряженные, и полярные, имеющие положительный или отрицательный заряд.
Под первичной структурой белка понимают порядок чередования аминокислотных остатков в полипептидной цепи. Первичная структура белка уникальна и детерминируется генами. К настоящему времени расшифрована первичная структура более тысячи белков из разных организмов, в том числе и человека.
Замена одного лишь аминокислотного остатка в полипептидной цепи может привести к аномальным явлениям. Примером тому служит замена в -цепи гемоглобина человека остатка глутамино-вой кислоты, занимающего шестое положение, на остаток валина. Результатом этого является тяжелое, передающееся по наследству заболевание — серповидноклеточная анемия.
Первичная структура белка предопределяет следующие уровни организации белковой молекулы.
Под вторичной структурой белка понимают способ укладки полипептидной цепи в упорядоченную структуру. По конфигурации выделяют следующие элементы вторичной структуры: -структура и -структура.
Третичная структура белка — это способ укладки полипептидной цепи в пространстве. Чтобы белок приобрел присущие ему функциональные свойства, полипептидная цепь должна определенным образом свернуться в пространстве, сформировав функционально активную структуру. Такая структура называется нативной. Несмотря на громадное число теоретически возможных для отдельной полипептидной цепи пространственных структур, сворачивание белка приводит к образованию единственной нативной конфигурации.
Белки с молекулярной массой более 100 кДа состоят, как правило, из нескольких полипептидных цепей со сравнительно небольшой молекулярной массой. Структура, состоящая из определенного числа полипептидных цепей, занимающих строго фиксированное положение относительно друг друга, вследствие чего белок обладает той или иной активностью, называется четвертичной структурой белка. Белок, обладающий четвертичной структурой, называется эпимолекулой или мультимером, а составляющие его полипептидные цепи — соответственно субъединицами или протомерами. Характерным свойством белков с четвертичной структурой является то, что отдельная субъединица не обладает биологической активностью.
Свойства белков
Белки — амфотерные электролиты. При определенном рН среды (изоэлектрическая точка) число положительных и отрицательных зарядов в молекуле белка одинаково. Это одна из основных констант белка. Белки в этой точке электронейтральны, а их вязкость и растворимость наименьшая. Способность белков снижать растворимость при достижении электронейтральности их молекул широко используется для выделения их из растворов, например в технологии получения белковых продуктов.
Белки связывают воду, т. е. проявляют гидрофильные свойства. При этом они набухают, увеличивается их масса и объем. Набухание белка сопровождается его частичным растворением. Гидрофильность отдельных белков зависит от их строения. Имеющиеся в составе и расположенные на поверхности белковой макромолекулы гидрофильные—СО—NH— (пептидная связь), аминные — NH2 и карбоксильные — СООН группы притягивают к себе молекулы воды, строго ориентируя их на поверхности молекулы.
Окружающая белковые глобулы гидратная (водная) оболочка препятствует агрегации, а следовательно, способствует устойчивости растворов белка и препятствует его осаждению.
Денатурация белков — сложный процесс, при котором под влиянием внешних факторов (температуры, механического воздействия, действия химических агентов и ряда других факторов) происходит изменение вторичной, третичной и четвертичной структуры белковой макромолекулы, т. е. ее нативной пространственной структуры. Первичная структура, а следовательно, и химический состав белка не меняются. При денатурации изменяются физические свойства белка, снижается растворимость, способность к гидратации, теряется его биологическая активность. Меняется форма белковой макромолекулы, происходит агрегирование. В то же время увеличивается активность некоторых химических групп, облегчается воздействие на белки протеолитических ферментов, а следовательно, он легче гидролизуется.
- Состав, структура и свойства ДНК и РНК
Биосинтез белка является завершающим этапом реализации генетической информации, закодированной в молекуле дезоксирибонуклеиновой кислоты (ДНК). Посредником в передаче генетической информации является матричная, или информационная, рибонуклеиновая кислота (мРНК). Биосинтез белка невозможен без участия еще двух видов РНК — транспортных (тРНК) и рибосомных (рРНК).
Понятие о нуклеиновых кислотах
Нуклеиновые кислоты — биополимеры, мономерами которых служат нуклеотиды, связанные между собой фосфодиэфирными связями.
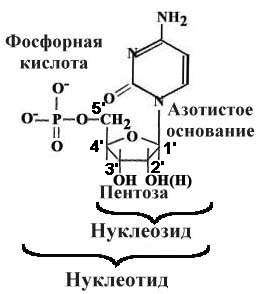
Нуклеотид = нуклеозид + фосфорная кислота = азотистое основание + пентоза + фосфорная кислота.
В РНК пентоза - рибоза. В ДНК - дезоксирибоза.
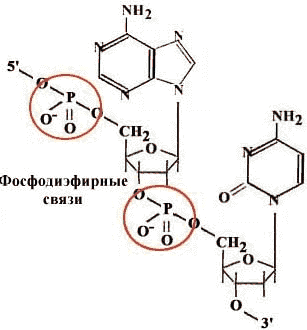
Нуклеотиды соединяются друг с другом в полимерную цепочку с помощью фосфодиэфирных связей. Азотистые основания не принимают участия в соединении нуклеотидов одной цепи.
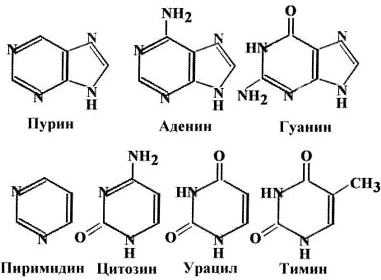
Существует два класса азотистых оснований. Пурины: аденин (А) и гуанин (Г) - содержат два гетероцикла. Пиримидины: тимин (Т), цитозин (Ц) и урацил (У) - содержат один гетероцикл.
В состав нуклеотидов входят следующие компоненты: углевод (рибоза в РНК, 2-дезоксирибоза в ДНК), азотистые основания (аденин, гуанин, цитозин и урацил в РНК; аденин, гуанин, цитозин и тимин в ДНК), фосфорная кислота. Примером трифосфатного нуклеотида может служить АТФ.
Транспортные РНК (тРНК) составляют 10% от всех РНК клетки. Основной функцией тРНК является перенос аминоацилов из цитоплазмы на рибосому, где происходит биосинтез белка. Каждая тРНК способна связывать только одну аминокислоту, поэтому конкретную тРНК называют по названию аминокислоты, которую она связывает.
Рибосомные РНК (рРНК) составляют основную массу клеточных (80—85%). В клетках эукариот существует несколько разных рРНК, отличающихся массой и структурой: две высокомолекулярные и две низкомолекулярные. Вместе с белками рРНК образуют рибосомы.
Рибосома представлена двумя субчастицами — большой и малой. Большая субчастица рибосомы состоит из одной высокомолекулярной рРНК, двух низкомолекулярных рРНК и 41 белка. В состав малой субчастицы рибосомы входит другая высокомолекулярная РНК и 31 белок. Кроме структурной, рРНК выполняют специфические функции в биосинтезе белка.
Принципы строения ДНК
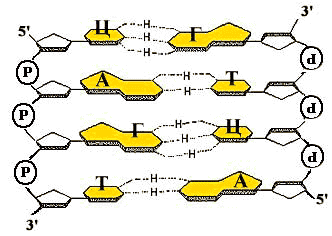 |
1. Нерегулярность. Существует регулярный сахарофосфатный остов, к которому присоединены азотистые основания. Их чередование нерегулярно.
2. Антипараллельность. ДНК состоит из двух полинуклеотидных цепей, ориентированных антипараллельно. 3`-конец одной расположен напротив 5`-конца другой.
3. Комплементарность (дополнительность). Каждому азотистому основанию одной цепи соответствует строго определенное азотистое основание другой цепи. Соответствие задается химией. Пурин и пиримидин в паре образуют водородные связи. В паре A-Т две водородные связи, в паре Г-Ц - три.
4. Наличие регулярной вторичной структуры. Две комплементарные, антипараллельно расположенные полинуклеотидные цепи образуют правые спирали с общей осью.
Виды РНК
| Виды РНК | Размер в нуклеотидах |
| gРНК - геномные РНК | 10000-100000 |
| mРНК - информационные (матричные) РНК | 100-100000 |
| tPHK - транспортные РНК | 70-90 |
| rРНК - рибосомные РНК | несколько дискретных классов от 100 до 500000 |
| sРНК - малые РНК | 100-300 |
Функции ДНК
1. ДНК является носителем генетической информации. Функция обеспечивается фактом существования генетического кода.
2. Воспроизведение и передача генетической информации в поколениях клеток и организмов. Функция обеспечивается процессом репликации.
3. Реализация генетической информации в виде белков, а также любых других соединений, образующихся с помощью белков-ферментов. Функция обеспечивается процессами транскрипции и трансляции.
Генетический код - это система записи информации о последовательности расположения аминокислот в белках с помощью последовательности расположения нуклеотидов в ДНК.
Поскольку ДНК непосредственного участия в синтезе белка не принимает, то код записывается на языке РНК. В РНК вместо тимина входит урацил.
3. Структура и кинетические характеристики ферментов
Ферментами, или энзимами (энзим от enzume—«в дрожжах», фермент от лат, fermentum — закваска), называют сложные биологические катализаторы белковой природы, изменяющие скорость химической реакции.
Раздел биохимии, изучающий биологические катализаторы белковой природы, называется этимологией. Круг вопросов, изучаемых энзимологией, весьма разнообразен. Он включает выделение и очистку ферментов с целью установления их состава и молекулярной структуры; изучение условий и скорости действия ферментов, а также влияния на них разнообразных физических и химических факторов.
Ферменты и катализаторы неорганической природы, подчиняясь общим законам катализа, имеют сходные признаки:
• катализируют только энергетически возможные реакции;
• не изменяют направление реакции;
• не расходуются в процессе реакции;
• не участвуют в образовании продуктов реакции.
Однако ферменты по ряду признаков отличаются от катализаторов неорганической природы. Главное отличие заключается в их химической природе, так как ферменты — белки.
В отличие от катализаторов неорганической природы ферменты «работают» в «мягких» условиях: при атмосферном давлении, при температуре 30—40 °С, при значении рН-среды близком к нейтральному. Скорость ферментативного катализа намного выше, чем небиологического. Единственная молекула фермента может катализировать от тысячи до миллиона молекул субстрата за 1 минуту. Такая скорость недостижима для катализаторов неорганической природы.
Ферменты обладают высокой специфичностью по отношению к субстрату; каждый фермент катализирует единственную реакцию либо группу реакций одного типа.
Ферменты являются катализаторами с регулируемой активностью, чего нельзя сказать о катализаторах иной природы. Это уникальное свойство ферментов позволяет изменять скорость превращения веществ в организме в зависимости от условий среды, т. е. приспосабливаться к действию различных факторов.
Существенным отличием ферментативных процессов является тот факт, что ферментативный процесс можно представить в виде цепи простых химических превращений вещества, четко запрограммированных во времени и в пространстве.
Строение ферментов
По строению ферменты делятся на простые (однокомпонентные) и сложные (двухкомпонентные). Простой фермент состоит только из белковой части; в состав сложного фермента входит белковая и небелковая составляющие. Иначе сложный фермент называют холоферментом. Белковую часть в его составе называют апоферментом, а небелковую — коферментом. Химическая природа коферментов была выяснена в 30-е гг. Оказалось, что роль некоторых коферментов играют витамины или вещества, построенные с участием витаминов В1, В2, В5, В6, В12, Н, Q и др. Особенностью сложных ферментов является то, что отдельно апофермент и кофермент не обладают каталитической активностью.
В составе как простого, так и сложного фермента, выделяют субстратный, аллостерический и каталитический центры.
Каталитический центр простого фермента представляет собой уникальное сочетание нескольких аминокислотных остатков, расположенных на разных участках полипептидной цепи. Образование каталитического центра происходит одновременно с формированием третичной структуры белковой молекулы фермента. Чаще всего в состав каталитического центра простого фермента входят остатки серина, цистеина, тирозина, гистидина, аргинина, аспа-рагиновой и глутаминовой кислот.
Субстратный центр простого фермента — это участок белковой молекулы фермента, который отвечает за связывание субстрата. Субстратный центр образно называют «якорной площадкой», где субстрат прикрепляется к ферменту за счет различных взаимодействий между определенными боковыми радикалами аминокислотных остатков и соответствующими группами молекулы субстрата. Субстрат с ферментом связывается посредством ионных взаимодействий, водородных связей; иногда субстрат и фермент связываются ковалентно. Гидрофобные взаимодействия также играют определенную роль при связывании субстрата с ферментом. В простых ферментах субстратный центр может совпадать с каталитическим; тогда говорят об активном центре фермента. Так, активный центр амилазы — фермента, гидролизующего -1,4-гликозидные связи в молекуле крахмала — представлен остатками гистидина, аспарагиновой кислоты и тирозина.
Аллостерический центр представляет собой участок молекулы фермента, в результате присоединения к которому какого-то низкомолекулярного вещества изменяется третичная структура белковой молекулы фермента, что влечет за собой изменение его активности. Аллостерический центр является регуляторным центром фермента.
В сложных ферментах роль каталитического центра выполняет кофермент, который связывается с апоферментом в определенном участке — кофермент связывающем домене. Понятия субстратного и аллостерического центров для сложного фермента и для простого аналогичны.
Механизм действия ферментов
Механизм действия простого и сложного ферментов одинаков, так как активные центры в их молекулах выполняют сходные функции.
Основы механизма действия ферментов были изучены в начале XX в. В 1902 г. английский химик А.Браун высказал предположение о том, что фермент, воздействуя на субстрат, должен образовать с ним промежуточный фермент — субстратный комплекс. Одновременно и независимо от А. Брауна это же предположение высказал французский ученый В. Анри. В 1913 г. Л. Михэлис и М. Ментэн подтвердили и развили представления о механизме действия ферментов, который можно представить в виде схемы:

где Е — фермент, S — субстрат, Р — продукт.
На первой стадии ферментативного катализа происходит образование фермент-субстратного комплекса, где фермент и субстрат могут быть связаны ионной, ковалентной или иной связью. Образование комплекса E-S происходит практически мгновенно.
На второй стадии субстрат под воздействием связанного с ним фермента видоизменяется и становится более доступным для соответствующей химической реакции. Эта стадия определяет скорость всего процесса.
На третьей стадии происходит химическая реакция, в результате которой образуется комплекс продукта реакции с ферментом.
Заключительным процессом является высвобождение продукта реакции из комплекса.
4. Витамины
Витамины — низкомолекулярные органические соединений различной химической природы, катализаторы, биорегуляторы процессов, протекающих в живом организме. Для нормальной жизнедеятельности человека витамины необходимы в небольших количествах, но так как в организме они не синтезируются в достаточном количестве, то должны поступать с пищей в качестве ее необходимого компонента. Отсутствие или недостаток в организме витаминов вызывает гиповитаминозы (болезни в результате длительного недостатка) и авитаминозы (болезни в результате отсутствия витаминов). При приеме витаминов в количествах, значительно превышающих физиологические нормы, могут развиваться гипервитаминозы. Людям еще в глубокой древности было известно, что отсутствие некоторых продуктов в пищевом рационе может быть причиной тяжелых заболевания (бери-бери, «куриной слепоты», цинги, рахита), но только в 1880 г. русским ученым Н. И. Луниным была экспериментальна доказана необходимость неизвестных в то время компонентом пищи для нормального функционирования организма. Свое название (витамины) они получили по предложению польского биохимика К.Функа (от лат. vita — жизнь). Сейчас известно свыше тридцати соединений, относящихся к витаминам. Различают собственно витамины и витаминоподобные соединения (полная незаменимость которых не всегда доказана). К последним относятся биофлавоноиды (витамины Р), пангамовая кислота (витамин B15), парааминобензойная кислота (витамин Н1) оротовая кислота (витамин В13), холин (витамин В4), инозич (витамин В8), метилметионинсульфонийхлорид (витамин U) липоевая кислота, карнитин (витамин В5). В отдельных продуктах содержатся провитамины, т. е. соединения, способные в организме превращаться в витамины. Например, -каротин переходит в витамин А, эргостеролы под действием ультрафиолетовых лучея в организме человека превращаются в витамин D.
Водорастворимые витамины. Витамин С (аскорбиновая кислота) необходим для нормальной жизнедеятельности человека; противоцинговый фактор, участвует в окислительно-восстановительных процессах, положительно действует на центральную нервную систему, повышает сопротивляемость человека к экстремальным воздействиям.
При недостатке аскорбиновой кислоты наблюдается характерная картина заболевания цингой: разрыхление и кровоизлияние десен, выпадение зубов.
Все необходимое количество витамина С человек получает с пищей. Основные источники витамина С: овощи, фрукты, ягоды. Содержание витамина С в свежем шиповнике 300—2000, черной, смородине 200—500, и капусте 30—70, молодом картофеле 20-30 мг %. При тепловой обработке пищи разрушается на 25—60%.
Витамин В1 (аневрин, тиамин) участвует в регулировании углеводного обмена. Недостаток вызывает нарушение в работе нервной системы, полиневрит (бери-бери). Необходим при ряде сердечно-сосудистых заболеваний. Основные источники: продукты из зерна (пшеничный и ржаной хлеб, хлеб из муки грубого помола), некоторые крупы (овсяная 0,5, ядрица 0,4 мг %), бобовых (горох 0,8, фасоль—0,5 мг %), мясопродукты (свинина 0,5 —0,6 мг %). Для увеличения содержания тиамина на мельзаводах проводят обогащение муки высшего и первого сорта синтетическим тиамином. Витамин В1 стоек к действию света, кислороду и к повышенным температурам в кислой среде. В щелочной среде легко разрушается, например, при добавлении в тесто щелочных разрыхлителей: соды, карбоната аммония.
Витамин B2 (рибофлавин) участвует в качестве кофермента в ферментных системах, катализирующих транспорт электронов в окислительно-восстановительных реакциях, которые протекают в живом организме. При недостатке рибофлавина возникают заболевания кожи, воспаление слизистой оболочки ротовой полости, появляются трещины в углах рта, развиваются заболевания кроветворной системы и желудочно-кишечного тракта. Источники витамина В2 (мг %): молоко — 0,15; творог — 0,3; сыр— 0,4, яйца — 0,4, хлеб — 0,1, ядрица — 0,2, мясо — 0,1—0,2, печень — 2,2, бобовые — 0,15, овощи и фрукты — 0,01— 0,06.
Витамин В6 (пиридоксин, адермин) участвует в синтезе и превращениях амино- и жирных кислот, входя в состав соответствующих ферментов. Необходим для нормальной деятельности нервной системы, органов кроветворения, печени. Недостаток витамина В6 вызывает дерматиты. Витамин В6 широко распространен в природе, основные его источники: мясные продукты (0,3—1 0,4 мг %), рыба (0,1 — 0,2 мг %), соя и фасоль (0,9 мг %), крупы (ядрица — 0,40 мг%, пшено — 0,52 мг%), картофель (0,30 мг%). Пиридоксин устойчив к повышенным температур рам, щелочам, кислотам, разрушается на свету. Некоторое количество пиридоксина поступает в организм в результате деятельности кишечной микрофлоры.
Витамин РР (ниацин, витамин B5). Под этим названием понимают два вещества, обладающих витаминной активностью: никотиновая кислота и ее амид (никотинамид). Ниацин является коферментом большой группы ферментов (дегидрогеназы), участвующих в окислительно-восстановительных реакциях, которые протекают в клетках. Никотинамидные коферменты играют важную роль в тканевом дыхании. При недостатке в организме витамина РР наблюдается вялость, быстрая утомляемость, бессонница, сердцебиение, пониженная сопротивляемость к инфекционным заболеваниям. При значительном недостатке развивается пеллагра (от ит. peilaagra — шершавая кожа) — тяжелое заболевание, приводящее к расстройству слизистой полости рта и желудка, появляются пятна на коже, нарушаются функции нервной и сердечно-сосудистой систем, психики.
Фолиевая кислота (витамин В9, фолацин, от лат. folium — лист) участвует в процессах кроветворения, перенося одноуглеродные радикалы, а также в синтезе амино- и нуклеиновых кислот, холина, пуриновых и пиримидиновых оснований. Фолиевая кислота широко распространена в природе, много ее содержится в зелени и овощах (мкг %): петрушке — 110, салате — 48, фасоли — 36, шпинате — 80, а также в печени — 240, почках — 56; твороге — 35—40, в хлебе — 16—27. Мало в молоке— 5 мкг %. Вырабатывается микрофлорой кишечника. При недостатке фолиевой кислоты наблюдаются нарушения кроветворения, пищеварительной системы, снижение сопротивляемости организма к заболеваниям.
Цианкобаламин (витамин В12) участвует в процессах кроветворения, превращениях аминокислот, биосинтезе нуклеиновый кислот. При недостатке витамина В12 появляется слабость, падает аппетит, развивается злокачественное малокровие, нарушается деятельность нервной системы. Для эффективного усвоения организмом человека витамина В12 необходим внутренний фактор - мукополисахарид слизистой желудка (внутренний фактор Косла), недостаток которою препятствует его всасыванию. Витамин В12 содержится в продуктах животного происхождения (мкг %): печени — 50—160, почках 20—30, рыбе 1—4, говядине — 2—6, сыре 1—2, молоке - 0.4.
Пантотеновая кислота (витамин В3) входит в состав ферментов биологического ацилирования, участвует в окислении и биосинтезе жирных кислот, липидов, в превращениях сахаров. Отсутствие пантотеновой кислоты в организме вызывает вялость онемение пальцев ног. Признаки гиповитаминоза наблюдаются редко. Пантотеновая кислота широко распространена в природе. Основные источники (мг %): печень и почки — 2,5—9, гречиха—1 2,6, рис — 1,7—2,1, овес - 2,5, яйца -- 1,4—2,7. Кулинарная обработка не приводит к существенному разрушению пантотеновой кислоты, но до 30% ее может переходить в воду при варке.
Жирорастворимые витамины. Витамин А (ретинол) участвует в биохимических процессах, связанных с деятельностью мембран клеток. При недостатке витамина А ухудшается зрение (ксерофтальмия — сухость роговых оболочек; «куриная» слепота). Замедляется рост молодого организма, особенно рост костей, наблюдается повреждение слизистых оболочек дыхательных путей, пищеварительных систем. Обнаружен только в продуктах животного происхождения, особенно его много в печени морских животных и рыб. В рыбьем жире — 15 мг %, печени трески — 4, .в сливочном масле 0,5, молоке — 0,025 мг %. Потребность человека в витамине А может быть удовлетворена и за счет растительной пищи, в которой содержатся его провитамины — каротины. Из молекулы -каротина образуется две молекулы витамина А. -Каротина больше всего в моркови — 9,0 мг %, красном перце — 2, помидорах — 1, сливочном масле — 0,2—0,4 мг %. Витамин А разрушается под действием света, кислорода воздуха, при кулинарной обработке (до 30 %). Поэтому плоды и овощи, содержащие провитамин А, необходимо обрабатывать быстро, не рекомендуется оставлять их надолго на воздухе в измельченном состоянии.
Кальциферол (витамин D) под этим термином понимают два соединения — эргокальциферол (D2) и холе кальциферол (D3). Регулирует содержание кальция и фосфора в крови, участвует в минерализации костей. Отсутствие приводит к развитию рахита у детей и разрежению костей (остеопороз) у взрослых. Следствие последнего — переломы костей, кариес зубов. Кальциферол содержится в продуктах животного происхождения (мкг %): рыбьем жире — 125, печени трески — 100, говяжьей печени — 2,5, яйцах — 2,2, молоке — 0,05, сливочном масле — 1,3—1,5. Потребность частично удовлетворяется за счет его образования в коже под влиянием ультрафиолетовых лучей из провитамина 7-дигидрохолестерина. Витамин D почти не разрушается при кулинарной обработке.
Токоферолы (витамин Е) предотвращают окисление ненасыщенных жирных кислот в липидах, влияют на биосинтез ферментов. При авитаминозе нарушаются функции размножения, сосудистая и нервная система. Распространены в растительных объектах, в первую очередь в маслах: в соевом — 115 мг %, хлопковом — 99, подсолнечником — 42 мг %; в хлебе — 2—4, ;крупах — 2—15 мг %. Витамин Е относительно устойчив к нагреванию, разрушается под влиянием ультрафиолетовых лучей.
- Обмен веществ
Живые организмы характеризуются рядом признаков, коренным образом отличающих их от неживых тел. Одним из таких признаков является обмен веществ — постоянно протекающий, самосовершающийся, саморегулирующий процесс обновления живых организмов. С прекращением обмена веществ прекращается жизнь. Обмен веществ включает в себя разнообразные физиологические, физические и химические процессы.
К физиологическим процессам относится поступление питательных веществ (белков, липидов, углеводов, минеральных веществ, воды, витаминов и др.) из окружающей среды и выделение продуктов жизнедеятельности организма. Физические процессы — это сорбция, всасывание, различные формы движения. К химическим процессам относятся распад питательных веществ и синтез необходимых организму соединений.
В химических процессах обмена веществ выделяют внешний и промежуточный виды обмена.
Внешний обмен — это внеклеточное превращение веществ на путях их поступления и выделения.
Промежуточный обмен — это превращения веществ внутри клеток. Процессы промежуточного обмена включают превращения компонентов пищи после их переваривания и всасывания. Именно промежуточный обмен изучает динамическая биохимия.
Промежуточный обмен веществ иначе называют метаболизмом. Метаболизм — это совокупность всех химических реакций в клетке. Вещества, образующиеся в ходе химических реакций, принято называть метаболитами. Число химических реакций в клетках организма человека огромно, но следует подчеркнуть, что все реакции протекают согласованно. Цепи химических реакций образуют метаболические пути или циклы, каждый из которых выполняет определенную функцию.
В метаболизме принято выделять два противоположных процесса — катаболизм и анаболизм.
Катаболизм (в переводе с греческого — вниз) — это процессы распада веществ, сопровождающиеся выделением энергии.
Анаболизм (в переводе с греческого — вверх) — процессы синтеза сложных молекул из более простых, сопровождающиеся потреблением энергии.
Катаболизм сопровождается освобождением энергии, которая может аккумулироваться в виде АТФ. При анаболических процессах происходит потребление энергии, которая освобождается при распаде АТФ до АДФ и фосфорной кислоты или АМФ и пирофосфорной кислоты. Следовательно, АТФ является сопрягающим энергетическим звеном катаболизма и анаболизма. Кроме АТФ связующим звеном могут служить специфические метаболические пути или циклы. Связующий путь (цикл), объединяющий пути распада и синтеза веществ, называется амфиболическим. Амфиболические пути связаны, как правило, с окислением веществ до углекислого газа и воды.
Основными функциями метаболизма являются:
• Распад структурных компонентов клетки;
• Аккумуляция энергии, извлекаемой при распаде химических веществ;
• Использование энергии для синтеза необходимых молекулярных компонентов и совершения работы.
Молодой растущий организм характеризуется преобладанием анаболических процессов над катаболическими. Это и понятно. Анаболические процессы обеспечивают рост организма, увеличение объема тканей и органов. Различия в средней скорости синтеза и распада веществ наиболее выражены сразу после рождения. К 17—19 годам в организме устанавливается динамическое равновесие между этими двумя фазами метаболизма. С этого возраста рост организма практически прекращается. К пожилому возрасту начинают преобладать катаболические процессы, что приводит к уменьшению содержания в организме ряда важнейших для жизнедеятельности веществ. Следствием этого является снижение силы мышц и функциональных возможностей внутренних органов.
Окружающая среда воздействует на организм порой разрушающе. В организме есть механизмы, способные поддерживать его в нормальном состоянии. Поддержание постоянного внутреннего состояния организма называется гомеостазом и является следствием метаболизма.
Распад белков
Главным путем распада белков в организме является ферментативный гидролиз, называемый протеолизом. Протеолиз белков протекает в любой клетке организма. Протеолитические ферменты локализованы в основном в лизосомах, незначительная часть ферментов, гидролизующих белки, есть в цитозоле клетки. Распад клеточных белков, катализируемый протеолитическими ферментами с различной специфичностью, приводит к образованию аминокислот, которые используются в этой же клетке или выделяются из нее в кровь. Но основным материалом для обновления клеточных белков служат аминокислоты, получаемые из белков пищи.
В желудочно-кишечном тракте локализованы протеолитические ферменты различной специфичности. В желудочном соке находится пепсин. Субстратом пепсина могут быть как нативные, так и денатурированные при термической обработке продуктов белки пищи. Пепсин быстро гидролизует в белках пептидные связи образованные карбоксильными группами ароматических аминокислот — фенилаланина, тирозина, триптофана. Медленнее пепсин гидролизует пептидные связи, образованные карбоксильными группами лейцина, аспарагиновой и глутаминовой кислот.
Протеолиз в кишечнике обеспечивает ряд ферментов: трипсин, химотрипсин, карбоксипептидазы А и В, дипептидазы и др.;
Трипсин гидролизует пептидные связи, образованные карбоксильными группами аргинина и лизина, химотрипсин — фенилаланина, тирозина и триптофана; действие этих ферментов приводит к более глубокому гидролизу белков по сравнению с гидролизом в желудке. Карбоксипептидаза А быстро отщепляет с С-конца образовавшихся олигопептидов аминокислоты с ароматическими или алифатическими боковыми радикалами. Карбоксипептидаза действует только на пептиды, имеющие на С-конце остатки аргинина или лизина.
Слизистая кишечника содержит группу аминопептидаз, которые при действии на полипептидные цепи поочередно освобождают N-концевые аминокислоты. Здесь же локализованы и дипептидазы, гидролизующие дипептиды.
Белки пищи при участии перечисленных ферментов гидролизуются до свободных аминокислот. На скорость гидролиза белков пищи указывает тот факт, что через 15 мин после приема человеком белка, содержащего меченые по азоту (15N) аминокислоты, изотоп 15N обнаруживается в крови. Максимальная концентрация аминокислот достигается через 30—50 мин после приема белка с пищей.
6. Углеводы
Углеводы — обширный класс органических соединений. В клетках живых организмов углеводы являются источниками и аккумуляторами энергии, в растениях (на их долю приходится до 90 % сухого вещества) и некоторых животных (до 20 % сухого вещества) выполняют роль опорного (скелетного) материала, входят в состав многих важнейших природных соединений, выступают в качестве регуляторов ряда важнейших биохимических реакций.
Строение, классификация и свойства углеводов
Все углеводы делят на две группы: простые и сложные. Простыми углеводами (моносахариды, монозы) называют углеводы, которые не способны гидролизоваться с образованием более простых соединений. Обычно их состав отвечает формуле Сn Н2n0n т. е. число атомов углерода равно числу атомов кислорода.
Сложные углеводы (полисахариды, полиозы) — углеводы, способные гидролизоваться на более простые. У них число атомов углерода не равно числу атомов кислорода. Сложные углеводы очень разнообразны по составу, молекулярной массе, а следовательно, и по свойствам. Их делят на две группы: низкомолекулярные (сахароподобные или олигосахариды) от греч. oligos — малый, немногочисленный и высокомолекулярные (несахароподобные полисахариды). Последние — соединения с большой молекулярной массой, в состав которых могут входить остатки сотен тысяч простых углеводов.
Молекулы простых углеводов — моноз — построены из неразветвленных углерод-углеродных цепей, содержащих различное число атомов углерода. В состав растений и животных входят главным образом монозы с 5 и 6 углеродными атомами — пентозы и гексозы. У атомов углерода расположены гидроксильные группы, а один из них окислен до альдегидной (альдозы) или кетонной (кетозы) группы:
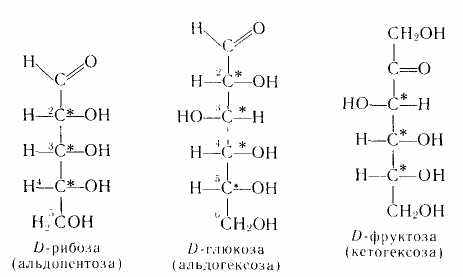
В водных растворах, в том числе в клетке, монозы из ациклических (альдегидокетоновых) форм переходят в циклические (фуранозные, пиранозные) и обратно. Этот процесс получил название динамической изомерии — таутомерии.
Молекулы полисахаридов построены из различного числа остатков моноз, которые образуются при гидролизе сложных углеводов. В зависимости от этого их делят на низкомолекулярные и высокомолекулярные полисахариды. Из первых особое значение имеют дисахариды, молекулы которых построены из двух одинаковых или разных остатков моноз. Одна из молекул моноз всегда участвует в построении молекулы дисахарида своим полуацетальным гидроксилом, другая — полуацетальным или одним из спиртовых гидроксилов. Если в образовании молекулы дисахарида монозы участвуют своими полуацетальными гидроксилами, образуется не-восстанавливающий дисахарид, во втором — восстанавливающий. Это одна из главных характеристик дисахаридов.
Мальтоза довольно широко распространена в природе, она содержится в проросшем зерне и особенно в больших количествах в солоде и солодовых экстрактах. Отсюда и ее название (от лат. maltum — солод). Образуется при неполном гидролизе крахмала разбавленными кислотами или амилолитическими ферментами, является одним из основных компонентов крахмальной патоки, широко используемой в пищевой промышленности. При гидролизе мальтозы образуются две молекулы глюкозы:
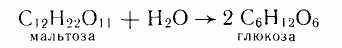
Этот процесс играет большую роль в пищевой технологии, например при брожении теста как источник сбраживаемых Сахаров.
Сахароза (тростниковый сахар, свекловичный сахар). При ее гидролизе образуются глюкоза и фруктоза:
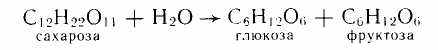
Следовательно, молекула сахарозы состоит из остатков глюкозы и фруктозы. В построении молекулы сахарозы глюкоза и фруктоза участвуют своими полуацетальными гидроксилами. Сахароза — невосстанавливающий сахар.
Лактозу получают из молочной сыворотки; отхода при производстве масла и сыра. В коровьем молоке содержится 4— 6 % лактозы. Отсюда и возникло ее название (от лат. lactum — молоко). Не участвует в спиртовом брожении, но под влиянием молочнокислых дрожжей гидролизуется с последующим сбраживанием образовавшихся продуктов в молочную кислоту (молочнокислое брожение).
Высокомолекулярные несахароподобные полисахариды построены из большого числа (до 6—10 тыс.) остатков моноз. Они делятся на гомополисахариды, построенные из молекул моносахаридов только одного вида (крахмал, гликоген, клетчатка), и гетерополисахариды, состоящие из остатков различных моносахаридов.
Крахмал (C6H10O10)n — резервный полисахарид, главный компонент зерна, картофеля и многих видов пищевого сырья. Наиболее важный по своей пищевой ценности и использованию в пищевой промышленности несахароподобный полисахарид.
Содержание крахмала в пищевом сырье определяется культурой, сортом, условиями произрастания, спелостью. Особенно крупные зерна у крахмала картофеля. О особенностей строении и размерен крахмальных зерен и, естественно, от состава крахмала зависят физико-химические свойства. Крахмал — смесь полимеров двух типов, построенный из остатков глюкопиранозы: амилозы и амилопектина. Их содержание в крахмале зависит от культуры и колеблется от 18 до 25 % амилазы и 75—82 % амилопектина.
Гликоген (животный крахмал) состоит из остатков глюкозы. Важный энергетический запасной материал животных (в печени до 10 %, мышцах 0,3—1 % гликогена) присутствует в некоторы растениях, например в зернах кукурузы. По своему строению напоминает амилопектин, но более разветвлен и его молекул имеет более компактную упаковку. Гликоген хорошо растворяется в горячей воде, но его растворы при охлаждении не образуют клейстер. Гидролизуется с образованием в качестве конечного продукта глюкозы.
Клетчатка — самый распространенный высокомолекулярный полимер (С6Н10О5)n. Это основной компонент и опорный материал клеточных стенок растений. Содержание клетчатки в волосках семян хлопчатника 98 %, древесине — 40—50, зернах пшеницы — 3, ржи и кукурузе — 2,2, сое — 3,8, подсолнечнике с плодовой оболочкой — до 15 %. Молекулы клетчатки с помощью водородных связей объединены в мицеллы (пучки), состоящие из параллельных цепей. Клетчатка нерастворима в воде и при обычных условиях не гидролизуется кислотами.
Гемицеллюлозы — это группа высокомолекулярных полисахаридов, образующих совместно с целлюлозой клеточные стенки растительных тканей. Присутствуют главным образом в периферииных оболочечных частях зерна, соломе, кукурузных початках, подсолнечной лузге. Содержание их зависит от сырья и достигает 40 % (кукурузные початки). В зернах пшеницы и ржи до 10% гемицеллюлоз. Гемицеллюлозы обычно имеют разветвленное строение; порядок расположения моноз внутри полимерной цепи неодинаков. Гемицеллюлозы широко применяют для получения разнообразных технических, медицинских, кормовых и пищевых продуктов, среди которых необходимо выделить агар и агарозу, ксилит. Гемицеллюлозы относят к группе пищевых волокон, необходимых для нормального пищеварения.
Пектиновые вещества — это группа высокомолекулярных полисахаридов, входящих в состав клеточных стенок и межклеточных образований растений совместно с целлюлозой, гемицеллюлозой, лигнином. Содержится в клеточном соке. Наибольшее количество пектиновых веществ находится в плодах и корнеплодах. Получают их из яблочных выжимок, свеклы, корзинок подсолнечника. Различают нерастворимые пектины (протопектины), которые входят в состав первичной клеточной стенки и межклеточного вещества, и растворимые, содержащиеся в клеточном соке. При созревании и хранении плодов нерастворимые формы пектина переходят в растворимые, с этим связано размягчение плодов при созревании и хранении. Переход нерастворимых форм в растворимые происходит при тепловой обработке растительного сырья, осветлении плодово-ягодных соков. Пектиновые вещества способны образовывать гели в присутствии кислоты и сахара при соблюдении определенных соотношений. На этом основано их использование в качестве студнеобразующего вещества в кондитерской и консервной промышленности для производства мармелада, пастилы, желе и джемов, а также в хлебопечении, сыроделии.
7. Липиды
Липидами называют сложную смесь органических соединений с близкими физико-химическими свойствами, которые содержатся в растениях, животных и микроорганизмах. Их общими признаками являются: нерастворимость в воде (гидрофобность) и хорошая растворимость в органических растворителях (бензине, диэтиловом эфире, хлороформе и др.), наличие в их молекулах длин-ноцепочечных углеводородных радикалов (R) и сложноэфирных
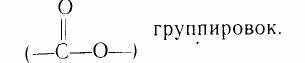
Строение и классификация липидов
По химическому строению липиды отличаются большим разнообразием. Молекулы их построены из различных структурных компонентов, в состав которых входят спирты и высокомолекулярные кислоты, а в состав отдельных групп липидов могут также входить остатки фосфорной кислоты, углеводов, азотистых оснований и другие компоненты, связанные между собой различными связями.
Липиды часто делят на две группы: простые и сложные.
Простые липиды. Молекула простых липидов не содержит атомов азота, фосфора, серы. К ним относят производные одноатомных (высших с 14—22 атомами углерода) карбоновых кислот и одно- и многоатомных спиртов (в первую очередь трехатомного спирта — глицерина). Наиболее важными и распространенными представителями простых липидов являются ацилглицерины. Широко распространены воски.
Ацилглицерины (глицериды) — сложные эфиры глицерина и высокомолекулярных карбоновых кислот. Они составляют основную массу липидов (иногда до 95—96 %) и именно их называют маслами и жирами. В состав жиров входят в основном триацилглицерины (триглицериды), но присутствуют ди- и моноацилглицерины:
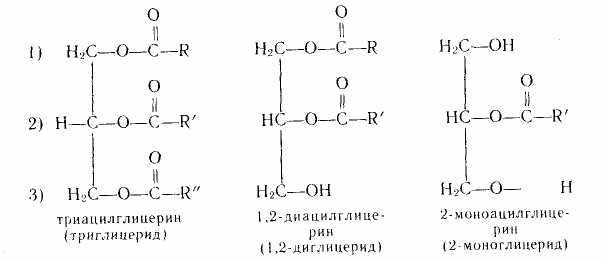
где R, R', R" — углеводородные радикалы.
Восками называют сложные эфиры высокомолекулярных одноосновных карбоновых кислот (С018 — С030) и одноосновных высокомолекулярных (с 18—30 атомами углерода) спиртов, входящие в состав липидов:
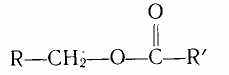
где R и R' — углеродные радикалы.
Они широко распространены в природе, покрывая тонким слоем листья, стебли, плоды растений, предохраняя их от смачивания водой, высыхания, действия микроорганизмов. Содержание их в зерне и плодах невелико. В оболочках семян подсолнечника содержится 0,2 % восков от массы оболочки, в семенах сои — 0,01, риса — 0,05 %.
Сложные липиды. Наиболее важная и распространенная группа сложных липидов — фосфолипиды. Молекула их построена из остатков спиртов, высокомолекулярных жирных кислот, фосфорной кислоты, азотистых оснований (чаще всего холина [НО—СН2—СН2—N(CH3)3] + ОН- и этаноламина НО—СН2— —СН2—NH2, аминокислот и некоторых других соединений.
Общая формула фосфолипидов (фосфатидов) имеет следующий вид:
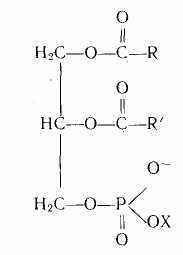
где R, R' — углеводные радикалы; X—Н; —СН2—CH2N(CH3)3; —СН2—СН2—NH3 и др.
Фосфолипиды — обязательный компонент клеток. Вместе с белками и углеводами фосфолипиды участвуют в построении мембран (перегородок) клеток и субклеточных структур (органелл), выполняя роль несущих конструкций мембран.
Фосфолипиды, выделенные в качестве побочных продуктов при получении масел, — хорошие эмульгаторы. Они применяются в хлебопекарной и кондитерской промышленностях, при производстве маргариновой продукции.
Стероиды широко распространены в природе, многочислены (до 20 тыс. соединений) и выполняют разнообразные функции в организме. Все стероиды — производные циклопентапергидрофенантрена; общий скелет стероидов имеет следующий вид (X - ОН, OR):
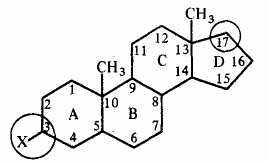
Из них выделим две группы: высокомолекулярные циклические спирты — стерины и их сложные эфиры. В молекуле стеринов у 3-го атома углерода (С-3) находится гидроксильная (—ОН) группа и у 17-го атома углерода (С-17) — разветвленная углерод-углеродная цепь (3-й и 17-й атомы обведены кружочками). Стерины нерастворимы в воде и хорошо растворимы в жирах. Несмотря на невысокое содержание, стерины и их производные играют исключительно важную роль в жизни живых организмов. В виде сложных комплексов с белками они входят в состав протоплазмы и мембран, регулируют обмен веществ в клетке.
Основные превращения липидов
Для глицеридов, составляющих основную массу масел и жиров, характерны следующие превращения: гидролиз, обмен остатков жирных кислот, входящих в их молекулы, окисление, гидрирование ненасыщенных ацилглицеринов. Гидролиз ацилглицеринов. Под влиянием фермента липазы, кислот или щелочей триацилглицерины гидролизуются с образованием ди-, затем моноглицеридов и в конечном итоге — жирных кислот и глицерина.
Переэтерификация. Ацилглицерины в присутствии катализаторов (метилат и этилат натрия, гидроксид натрия, ферменты) способны к обмену (миграции) остатков жирных кислот. Этот процесс получил название переэтерификации. В результате переэтерификации меняется ацилглицериновый состав жира, а следовательно, меняются и их физико-химические свойства.
Обмен кислотных остатков может происходить между молекулами ацилглицеринов (межмолекулярная переэтерификация) и в пределах одной молекулы (внутримолекулярная переэтерификация).
Переэтерификация открывает большие возможности для изменения свойств жира (температуры его плавления, затвердевания, пластичности), т. е. позволяют получать жиры с заданными для пищевой технологии физико-химическими свойствами без изменения их жирокислотного состава.
Гидрогенизация. Остатки ненасыщенных жирных кислот в молекулах ацилглицеринов присоединяют водород. Этот процесс получил название гидрогенизации масел и жиров. С его помощью целенаправленно изменяется жирнокислотный, а следовательно, и ацилглицериновый состав масел, повышается их температура плавления, твердость, меняется пластичность. Они становятся, более стойкими к окислению.
Пищевая порча жиров. При хранении растительные масла и животные жиры, жиросодержащие продукты (масличные семена, мука, крупа, кондитерские изделия, рыба, мясные продукты) под влиянием кислорода воздуха, света, ферментов и т. д. постепенно меняют свои, присущие им свойства, в первую очередь органолептические. Характер изменений вкуса и запаха зависит от вида жира, особенностей протекающих в нем процессов и связан с преимущественным накоплением в продукте тех или иных химических веществ, имеющих разную природу. Одновременно в жирах накапливаются вредные для организма человека продукты окисления пипидов. В результате снижается их пищевая и физиологическая ценность и они могут оказаться непригодными для употребления, т. е. произошла пищевая порча жира, его прогоркание.
Прогоркание жиров и жиросодержащих продуктов — результат сложных химических и биохимических процессов, протекающих в липидном комплексе. В зависимости от характера основного процесса, протекающего при этом, различают гидролитическое и окислительное прогоркание. Каждый из них может быть разделен на автокаталитическое (неферментативное) и ферментативное (биохимическое) прогоркание.
Список использованной литературы
- Д.Г. Кнорре, С.Д. Мызина. Биологическая химия. - М.: Высшая школа 2002г. (библ. ИнЕУ)
- А. Ленинджер. Биохимия. М. Мир 1974 г. (библ. ИнЕУ)
- Ю.Б.Филиппович. Основы биохимии. М.: Высшая школа, 1985
- К.А. Арыстанова, З.А. Мустафина и Э.Ж. Алимкулова. Практикум по биохимии. Астана: Казахский аграрный университет им. С. Сейфуллина, 2003.
- В.Л. Кретович. Биохимия растений. -М.: Высшая школа,1986
- Вальтер О.А., Пиневич Л.М., Варасова Н.Н. Практикум по физиологии растений с основами биохимии. М.Л.: Сельхозгиз, 1957 г.
- А. Паттон Энергетика и кинетика биохимических процессов. М. Мир. 1968 г.
- М.П. Шерстнев, О.С. Комаров Химия и биология нуклеиновых кислот М. Просвещение 1990 г. (библ. ИнЕУ)
- Некрасов В.В. Руководство к малому практикуму по органической химии. -М.Л.: Химия, 1964 г.
- Плешков Б.П. Практикум по биохимии растений. -М.: Колос,1968 г.
- В.И. Добрынина. Учебник биологической химии. -М.: Госиздатметлит, 1963 г.