
Структурно-функциональные особенности запасных и защитных белков растений и их использование в генетических исследованиях
| Вид материала | Автореферат |
- Структурные и функциональные свойства цитокинин-связывающих белков растений Arabidopsis, 249.12kb.
- Педагогическая пресса в россии во второй половине XIX в.: Генезис, предметно-тематические, 302.96kb.
- Рабочая учебная программа по нормальной физиологии Электив «Структурно-функциональные, 112.81kb.
- Нп «сибирская ассоциация консультантов», 169.41kb.
- Календарно-тематический план практических занятий по биологии для студентов медико-биологического, 71.39kb.
- Названия экзотических растений в английском и русском языках (структурно-словообразовательный, 378.31kb.
- Программа 9 16 ноября 2010 г г. Москва Место проведения конференции: Учреждение Российской, 265.18kb.
- Добриева Зейнаб Израиловна структурно-семантические и функциональные особенности приставочных, 2630.63kb.
- Структурно-семантические и функциональные характеристики английского лингвокультурного, 562.68kb.
- Программа для поступающих в нввкус по специальности «Психология», 284.28kb.
6. Исследование полиморфизма белков семян огурца
Выше было показано, как исследование полиморфизма проламинов злаков (по N-концевым последовательностям) может быть использовано для решения проблем структуры глиадинкодирующих кластеров генов и филогении пшеницы. Однако эти исследования имеют и практическое значение – для решения вопросов частной генетики и селекции важнейших сельскохозяйственных культур, таких как идентификация генотипов, определение сортовой чистоты, выявление гибридов. В то же время подобные маркеры у овощных культур, в частности, тыквенных, отсутствовали. Нами были проведены исследования по разработке методов электрофоретического анализа белков семян тыквенных для использования их в экспериментах по частной генетике и в сортовом семенном контроле данных культур. Исследованию подвергали фракции альбуминов, глобулинов и основного глобулина семян – кукурбитина. В качестве метода выявления полиморфизма белков использовали различные электрофоретические системы.
Исследование альбуминов семян огурца путем электрофореза в ПААГ в присутствии ДДС-Na выявил многокомпонентность электрофоретических спектров и незначительное количество сортоспецифических компонентов, что затрудняет использование данной системы для генетического анализа и массового применения в сортовом семенном контроле огурца.
Электрофоретический анализ глобулинов в этой же электрофоретической системе позволил обнаружить межлинейные различия по минорным компонентам в молекулярно-весовом диапазоне от 49 кДа и выше. Межсортовые различия как между зарубежными, так и между отечественными сортами огурца не были выявлены. Таким образом, на исследованном материале электрофорез глобулинов также не выявил сортоспецифичности спектров.
Исследование полиморфизма кукурбитина – основного запасного глобулина - с помощью электрофореза в ПААГ в присутствии ДДС-Na показало, что в использованной электрофоретической системе фракция кукурбитина у исследованных образцов огурца является инвариантной. У двадцати двух исследованных сортов и линий вариабельным оказался лишь компонент с молекулярной массой 36 кДа, который либо присутствовал, либо отсутствовал в спектре. Таким образом, на изученном материале удалось выделить лишь 2 типа электрофоретических спектров кукурбитина огурца. Суммируя полученные данные, можно сделать вывод о том, что альбумины и глобулины исследованных образцов огурца выявили низкий межсортовой полиморфизм, что затрудняет использование этих белков для идентификации сортов и установления процента гибридности семян в семенном контроле. Идентификация генетически отдаленных форм возможна.
7. Исследование полиморфизма белков семян тыквы
Изучение гетерогенности белков семян проводили на районированных сортах тыквы отечественной селекции. В коллекцию входили сорта Cucurbita pepo, Cucurbita maxima и Cucurbita moschata. Анализ альбуминов и глобулинов семян тыкв выявил их большую в сравнении с огурцом гетерогенность и специфичность. Более полиморфной у представителей рода Cucurbita оказалась и фракция кукурбитина. При разделении кукурбитина в денатурирующих условиях в электрофоретическом спектре было выделено 3 зоны: α-, β- и γ- (рис. 11). Вариабельные компоненты находились в α-, и γ-зонах. Наиболее полиморфной оказалась низкомолекулярная γ-зона (молекулярная масса субъединиц 21-26 кДа), где в сортовых спектрах обнаруживалось от 4 до 7 компонентов (рис. 11, 12).

Рис. 11. Электрофореграммы кукурбитина семян сортов C. pepo, C. maxima и C. moschata.
Электрофоретический анализ белков гибридных семян и их родительских форм показал, что субъединицы кукурбитина наследуются кодоминантно, что делает возможным идентифицировать семена гибридов, которые возникают при естественном переопылении разновидностей C. pepo (рис. 12в).
Р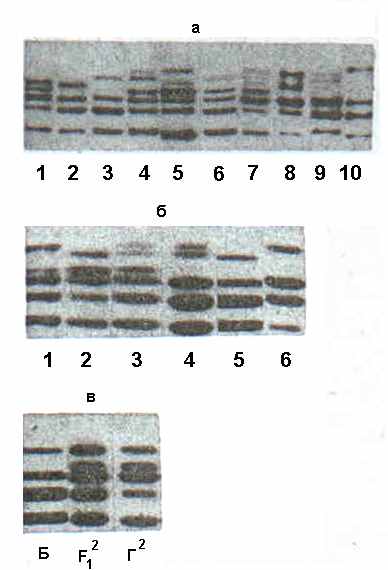 ис. 12. Электрофореграммы субъединиц кукурбитина в ПААГ в присутствии ДДС-Na, γ-зона: (а) сорта C. pepo (1 – Якорь, 2 – Белоплодные, 3 – Сотэ 38, 4 – Грибовские 37; 5 – Одесские 52; 6 – Греческие 110; 7 – Грибовская кустовая; 8 – Украинская многоплодная); 9 – C. maxima (Грибовская зимняя); 10 – C. moschata (Перехватка); (б) биотипы сорта Грибовские 37; (в) сорт Белые 13 (C. p. var. melopepo) – Б, сорт Грибовские 37 (биотип 2, C. p. var. giraumons) – Г2, гибрид Б х Г2-F12.
ис. 12. Электрофореграммы субъединиц кукурбитина в ПААГ в присутствии ДДС-Na, γ-зона: (а) сорта C. pepo (1 – Якорь, 2 – Белоплодные, 3 – Сотэ 38, 4 – Грибовские 37; 5 – Одесские 52; 6 – Греческие 110; 7 – Грибовская кустовая; 8 – Украинская многоплодная); 9 – C. maxima (Грибовская зимняя); 10 – C. moschata (Перехватка); (б) биотипы сорта Грибовские 37; (в) сорт Белые 13 (C. p. var. melopepo) – Б, сорт Грибовские 37 (биотип 2, C. p. var. giraumons) – Г2, гибрид Б х Г2-F12.
Таким образом, по электрофоретическим спектрам кукурбитина нами был выявлен межвидовой, внутривидовой и внутрисортовой полиморфизм, что открывает возможности использования метода электрофореза кукурбитина для экспресс-анализа сортовой чистоты образцов тыквы обыкновенной. Меньшая сортовая специфичность кукурбитина у крупноплодной и мускатной тыквы позволяет идентифицировать лишь группы сортов, что требует разработки и применения альтернативных систем. Таковой для представителей рода Cucurbita может служить система электрофореза альбуминов в кислой среде (рН 3,1).

Рис. 13. Электрофореграммы альбуминов семян сортов C. pepo, C. maxima, C. moschata.
В этой системе альбумины семян разделялись на 40 вариабельных компонентов (рис. 13). Были выделены компоненты, специфичные для каждого из изученных видов, для разновидностей «перехватка» (№56-58) и «продолговатая» (№59-60) мускатной тыквы. Различные сочетания альбуминов обеспечивают сортоспецифичность спектров всех изученных сортов. Исключение, как и по кукурбитину, составляли кабачки Зебра (№28) и Цукеша (№29), что делает возможным применение данной системы электрофореза в семенном контроле наряду с электрофорезом кукурбитина.
8. Генетический контроль компонентного состава кукурбитина
Одной из наиболее серьезных форм биологического засорения образцов тыквы является присутствие в нем гибридов от переопыления между разновидностями тыквы обыкновенной в коллекционных и селекционных питомниках. Выявление подобного типа засорения при грунтконтроле осложняется тем, что основной апробационный признак - форма семенника - у тыкв определяется комплементарным действием генов. В связи с этим нами была исследована возможность выявления гибридов в посевном материале по электрофоретическим спектрам белков семян на основе изучения генетического контроля отдельных компонентов кукурбитина.
Был проведен гибридологический анализ состава субъединиц кукурбитина в гибридной комбинации патиссон Белые 13 х кабачок Грибовские 37. Сорт кабачков Грибовские 37 обладает генотипом, типичным для стародавних сортов. В результате было установлено, что, во-первых, наследование вариабельных компонентов кукурбитина в спектрах сортов Белые 13 и Грибовские 37 осуществляется тремя локусами (А, В и С). Генотипы родительских сортов содержат различные аллельные варианты локусов А, В и С. Во-вторых, аллели одного локуса контролируют отдельные компоненты кукурбитина, различающиеся по молекулярной массе, в отличие от блоков компонентов, выделенных в качестве единицы наследования у проламинов злаков.
Анализ расщепления одновременно по трем кукурбитинкодирующим локусам свидетельствует о том, что они не сцеплены между собой: соотношение 18 классов, обнаруженных в F2, полностью соответствует теоретически ожидаемому расщеплению (1:2:1):(1:2:1): (3:1) при χ2факт.=12,4, χ2теор. = 27,6.
Для формализации данных гибридологического анализа нами предложена классификация кукурбитина, позволяющая выражать электрофореграммы в виде формул. Для этого в строку последовательно заносится название белковой системы, буквенное обозначение локуса и порядковый номер идентифицированного компонента, кодируемого соответствующим аллелем. Например, электрофореграмму сорта Грибовские 37 (рис. 12г) можно записать как CbnA2 B1 C1. В генетических формулах гетерозигот по кукурбитину аллели одного локуса разделяются запятой. Например, формула гибрида F1 от скрещивания Белые 13 х Грибовские 37 (рис. 12б,в) будет CbnA1,2 B1,2 C1,0. В сокращенном виде формула сорта Грибовские 37 будет 2.1.1., а упомянутого выше гибрида F1 – 1,2.1,2.1,0.
Возможность идентификации гибридов была продемонстрирована также на комбинации скрещивания, включающей сорт кабачков современной селекции Якорь и патиссон Белые 13. Кроме того, была показана возможность идентификации гибридов еще одного варианта скрещивания: тыква х кабачок (на примере сортов Миндальная 35 х Сотэ 38).
Изучение коллекции сортов тыкв с применением гибридологического анализа, а также методов идентификации аллелей сравнительным анализом электрофореграмм разных биотипов гетерогенного сорта и с использованием эталонного сорта позволило составить каталог аллельных вариантов компонентов кукурбитина, контролируемых локусами Cbn A, Cbn B, Cbn C, Cbn D. По локусам А, С, D обнаружены пары аллелей, по локусу В – серия, состоящая из 7 множественных аллелей (рис. 14).
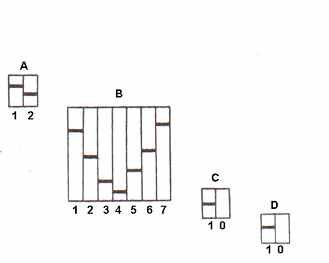
Рис. 14. Каталог аллельных вариантов компонентов электрофоретического спектра кукурбитина, контролируемых локусами Cbn A, Cbn B, Cbn C, Cbn D.
Каталог систематизирует полученные данные по генетическому контролю компонентов кукурбитина, делает возможным предсказание фенотипических классов по кукурбитину при гибридизации.
При анализе характера распределения аллелей кукурбитинкодирующих локусов у сортов различных видов тыкв было установлено, что наибольшая генетическая изменчивость по четырем локусам наблюдается у вида C. pepo – H=0,116, несколько ниже она у C. moschata - H=0,056 и C. maxima – H=0,041. Полученные результаты совпадают с существующией точкой зрения, что тыква обыкновенная является наиболее полиморфной из трех культивируемых в нашей стране видов.
Исследование полиморфизма кукурбитина на внутривидовом уровне показало, что электрофоретические спектры кукурбитина, как правило, сортоспецифичны. Однако при этом выделяются группы сортов, имеющих в генотипе одинаковый набор аллелей кукурбитинкодирующих локусов, различие между подобными образцами выражаются в интенсивности электрофоретически выявляемых компонентов.
Путем параллельной оценки чистосортности партии семян суперэлиты патиссонов Белые 13, выращенной на Симферопольской ОБОС, двумя методами была продемонстрирована высокая эффективность предложенного метода (по электрофоретическим спектрам куккрбитина) по сравнению с традиционно используемым грунтконтролем.
Глава II. Защитные белки
В настоящем разделе диссертации излагаются результаты исследования защитных PR-белков, образующихся в растениях в ответ на инфицирование, для выяснения механизмов устойчивости к патогенам. Цель работы состояла в выявлении и изучении свойств PR-белков у растений рода Nicotiana и Lycopersicon с разными генами устойчивости при вирусной инфекции и перекрестной защите.
1. Обнаружение PR-белков в листьях разных видов табака при вирусной инфекции
Предварительно было установлено, что механическое повреждение листьев табака индуцирует образование новых белков, Rf которых составили 0,84 и 0,67. Такие растения были использованы в качестве контроля при изучении влияния вирусной инфекции на спектр кислоторастворимых белков листьев. Изучение состава белков листьев гибридного табака Терновского в динамике (на 3, 7, 10 и 14 день после заражения) показало, что содержание PR-белков наибольшее на 10 день после заражения, после чего (на 14 день) их содержание падает, что сопровождается некротизацией листьев. Исследование накопления PR-белков в разных участках ткани зараженного листа, а также в других частях растения показало, что PR-белки преимущественно накапливаются в непосредственной близости от места инокуляции вирусом. При удалении от места инфицирования их содержание падает: так, на расстоянии 2 см от некроза их содержание падает почти в 2 раза. В верхних неинокулированных листьях PR-белки обнаруживаются лишь в следовых количествах. Был проанализирован спектр PR-белков у разных видов и сортов табака и выявлена видоспецифичность набора PR-белков (табл. 4). Это позволяет использовать их в качестве генетических маркеров как при исследовании происхождения разных видов табака, так и при анализе устойчивости растений табака к вирусным заболеваниям. В то же время межсортовых различий по электрофоретическим спектрам PR-белков выявлено не было.
Исследование кислоторастворимых белков листьев табака Терновского при системной реакции на заражение ВОМ-1 показало, что в отличие от других исследованных видов табака, у гибрида PR-белки образуются и при системном заражении (табл. 4).
Таблица 5.
PR-белки, выявленные у разных видов и сортов Nicotiana при вирусной инфекции
| Вид, сорт | Ген | Реакция на ВТМ | PR-белки и их молекулярные массы, кДа | ||||||||
| b0 | b1 | b1 | b2 | b3 | b4 | b5 | b6 | b7 | |||
| 16 | 16,6 | 14 | 16,2 | 16 | - | 34 | 35 | 35 | |||
| N. glutinosa | N | сверхч. | - | - | + | - | - | - | - | - | - |
| N. rustica | N | сверхч. | - | - | - | + | - | - | + | + | + |
| N. sylvestris | N´ | сверхч. | + | + | - | - | + | - | - | - | - |
| системн. | - | - | - | - | - | - | - | - | - | ||
| N. tabacum сорт Samsun EN | N´ | сверхч. | - | + | - | + | + | + | - | - | - |
| системн. | - | - | - | - | - | - | - | - | - | ||
| N. tabacum сорт Java | N´ | сверхч. | - | + | - | + | + | + | - | - | - |
| системн. | - | - | - | - | - | - | - | - | - | ||
| Гибридный табак | N | сверхч. | - | + | - | + | + | + | + | + | + |
| системн.* | - | + | - | + | + | + | + | + | + | ||
*заражение ВОМ-I; сверхч.- сверхчувствительная реакция; системн. – системная реакция.