
Экология зоопланктона разнотипных водоемов бассейна Верхней Волги
| Вид материала | Автореферат диссертации |
- Зоопланктон разнотипных водных экосистем бассейна верхней оби, 420.52kb.
- Республика Марий Эл расположена в центре европейской части Российской Федерации,, 938.98kb.
- Партийно-государственная политика по формированию инженерно-технической интеллигенции, 390.4kb.
- Экология водоемов зоны техногенной радиационной аномалии на южном урале 03. 00., 1110.67kb.
- Славяне Введение, 467.33kb.
- Эколого-гидрологические исследования водоемов, 425.38kb.
- Культура Руси в период, 138.66kb.
- Промышленная экология методы и способы защиты воздушного бассейна, 721.61kb.
- Тульский Государственный Университет кафедра"Истории и культурологии" Контрольно-курсовая, 141.72kb.
- Эколого-фаунистический анализ паразитофауны рыб вислинского залива, рек преголи и прохладной, 419.14kb.
Разграничение экологических последствий антропогенной и естественной ацидификации представляет достаточно сложную проблему. В бассейне Верхней Волги только гумифицированные озера испытывают влияние поступления органических анионов с водосбора, светловодные закислены преимущественно кислотными атмосферными осадками (Комов, Лазарева, 1994). Сравнивая светловодные и гумифицированные водоемы с рН воды <5.3, можно представить как развиваются изменения в биоте, если озера закислены только антропогенно и если причин закисления две. Сообщества гидробионтов гумифицированных озер с рН воды 5.5-6.0 характеризуют переходные, а с рН >6 – природно-закисленные биотопы.
При закислении водоемов формируются два типа воздействий на сообщества: «прямые» и «косвенные», которые при взаимодействии могут усиливать или ослаблять друг друга. «Прямые» эффекты вызваны нарушением у гидробионтов в кислой среде кислотно-щелочного равновесия, ионной и осмотической регуляции, дыхания, обмена кальция, сахаров и аминокислот, а также усилением биоаккумуляции ксенобиотиков (Hg, Mn, Cd, Cu, Ni) (Виноградов и др., 1979; Stenson, Eriksson, 1989; Locke, 1991; Haines et al. 1994, 1995; Brakke et al., 1994; Yan et al., 1996; Степанова,Комов, 1996, 1997). «Косвенное» влияние закисления вод на биоту связывают с изменением трофических взаимодействий в обедненных видами сообществах, с трансформацией биотопической структуры экосистем озер из-за исчезновения макрофитов, развития матов водорослей или мхов и изменения глубины эвфотной зоны, а также с редукцией запаса и/или доступности для биоты таких важных биогенных элементов, как азот, фосфор и неорганический углерод (Grahn et al., 1974; Broberg, Persson, 1984; Rudd et al., 1988; Turner et al., 1991; Fairchield, Sherman, 1992; Gahnstrom et al., 1993; Sherman, Fairchield, 1994; Schindler, 1994; Brakke et al., 1994). В сообществе зоопланктона наиболее чувствительны к прямому токсическому действию закисления рачки сем. Daphniidae, наименее – кладоцеры сем. Macrotricidae, циклопы и коловратки (наш анализ по: Brett, 1989; Locke, 1991). Даже самые чувствительные виды (Daphnia sp.) в условиях водоема элиминируются при более высоком уровне рН, чем в эксперименте (Muniz, 1991; Лазарева, 1995), что свидетельствует о значительном влиянии косвенных факторов на развитие их популяций. Биоаккумуляция ртути в тканях рыб в закисленных озерах усиливается при высокой биомассе рачкового зоопланктона (Комов, 1999). Таким образом, «прямое» влияние накопления ксенобиотика усиливается посредством «косвенных» взаимодействий в трофической сети водоема.
По суммарным спискам зоопланктона нейтральные (рН>6.5) и ацидные (рН<5) водоемы почти не различаются, индексы сходства Шимкевича-Симпсона >70% (Лазарева, 1992а, 1994). Рассуждения исследователей об изменении состава зоопланктона под влиянием закисления, как правило, относятся к набору доминантных форм (Салазкин, 1976; Андроникова, 1992, 1996; Иванова, 1997). Почти все виды летнего зоопланктона встречены в диапазоне рН 4.1-9.1 и РОВ 1.9-62.7 мгС/л (Лазарева, 1998). Однако их высокая численность четко приурочена к определенному интервалу градиента факторов среды (абсцисса центра доминирования на рис. 3). Близость центров доминирования определяет группы доминантов, наиболее вероятные для данного диапазона условий. Первые четыре вида образуют доминантный комплекс (доминантный тип С по: Lazareva, 1995), характерный для гумифицированных (РОВ >10 мг С/л) озер с рН воды >6.5 (встречаемость 67 и 88% соответственно). Виды с пятого по седьмой формируют доминантный тип В, свойственный переходным озерам с рН воды >5.5 и РОВ >10 мг С/л (встречаемость 68 и 87%), а последние пять видов – доминантный тип А, обычный для водоемов с рН <5.3 и широким диапазоном РОВ (встречаемость >48%). Для доминантного типа А зоопланктона характерны две отличительные черты: высокая доля в сообществе копепод Eudiaptomus graciloides (40-55% биомассы зоопланктона) и формирование «бидоминантных» (копепода + кладоцера) комплексов, в которых два вида дают до 90% средней за сезон Вобщ зоопланктона (Лазарева, 1994). Уровень доминирования по Симпсону в них достигает 0.66-0.93 (Лазарева, 1993). Преобладание (>70% Вобщ) в планктоне закисленных озер северо-запада России немногих видов (чаще кладоцер) отмечают и другие исследователи (Салазкин, 1976; Иванова, 1997; Вандыш, 2002).
Одним из косвенных методов изучения сукцессий служит установление сукцессионных рядов на основании анализа пространственных (Миркин, Розенберг, 1978; Шитиков и др., 2005). Если пространственный градиент фактора среды выступает одновременно индикатором возраста или сукцессионной стадии, то результирующая ординационная схема может быть названа экохроноклином (Миркин, Наумова, 1983). Экохроноклины изменения встречаемости и доминирования видов зоопланктона в озерах по градиенту рН воды позволили отчетливо проследить смены зоопланктона при закислении озер (Лазарева, 1998). Изменения зоопланктона, вызванные гумификацией (РОВ), выражены менее четко, поскольку часть видов безразлична к содержанию РОВ. Смены видов по градиентам рН воды и содержания РОВ в данном случае отражают сукцессию сообщества: аллогенную, вызванную антропогенной ацидификацией, и аутогенную, связанную с гумификацией и природным закислением вод озер. Последовательность замещения видов зоопланктона по мере увеличения уровня гумификации, фактически противоположна, наблюдаемой при снижении рН воды (рис. 3). Вероятно, поэтому в антропогенно закисленных гумифицированных озерах часто наблюдают менее выраженные изменения в зоопланктоне, чем в светловодных (Sarvala, Halsinaho, 1990; Lazareva, 1995; Лазарева, 1996), что позволяет говорить о том, что высокая концентрация РОВ «смягчает» действие закисления (Schindler et al., 1991).
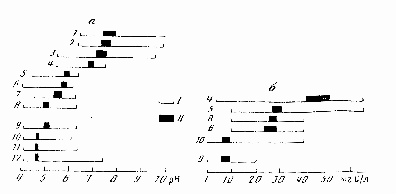
Рис. 3. Экохроноклины доминирования видов озерного зоопланктона по градиентам рН воды (а) и РОВ (б) (по: Лазарева, 1998).
I – эмпирический интервал доминирования (относительная численность вида >10%), II – доверительный интервал 95% экологических центров доминирования видов. 1 – Daphnia cristata, 2 – Mesocyclops leuckarti, 3 – Keratella cochlearis, 4 – Thermocyclops oithonoides, 5 – Asplanchna priodonta, 6 – Holopedium gibberum, 7 – Eudiaptomus gracilis, 8 – Ceriodaphnia quadrangula, 9 – Bosmina obtusirostris, 10 – Diaphanosoma brachyurum, 11 – Polyarthra vulgaris, 12 – Eudiaptomus graciloides.
По мере снижения рН воды изменяется распределение обилия между видами зоопланктона. В закисленных водоемах доминируют немногие виды и S-образных кривые «разнообразия-доминирования» сменяются прямыми (Лазарева, 1993). Как следствие увеличивается доля (до >80% Sобщ) малочисленных видов. Это служит основной причиной неравномерного накопления видов в суммарных списках зоопланктона ацидных и нейтральных водоемов, а также уменьшения числа видов в единичной пробе (Sпр) и показателей разнообразия (Hb). В антропогенно закисленных озерах (типы 1 и 2) Sпр = 4-18, Hb = 1.33±0.43, в нейтральных (тип 3) – Sпр = 11-23, Hb = 1.51-3.00 (Лазарева, 1993; Lazareva, 1995). Высокая концентрация РОВ в природно закисленных водоемах (тип 4) тоже приводит к увеличению уровня доминирования и снижению богатства и разнообразия зоопланктона (Hb = 0.50±0.10) (Лазарева, 1993). Наибольшее влияние на таксономическую структуру зоопланктона оказывают факторы, характеризующие уровень закисления (рН, ANC) и трофический статус (Ptot, Хл. а) (табл. 3). Снижение индекса Шеннона для зоо- и фитопланктона при уменьшении рН с 7 до 4 выявлено для многих озер Европейской России (Ivanova et a., 1993; Андроникова, 1996; Иванова, 1997; Моисеенко и др., 1999; Вандыш, 2002), а также Северной Америки (Siegfried et al., 1989). Уменьшение видового разнообразия можно отнести к наиболее общим откликам зоопланктона на закисление вод любой природы.
Таблица 3. Факторы, наиболее сильно влияющие на характеристики зоопланктона малых озер (по: Лазарева, 1993, 1995).
| Показатель | Фактор | R | R2,% | F |
| Sпр | рН | 0.55 | 30 | 21.9 |
| (37 озер) | Ptot | 0.53 | 15 | 9.6 |
| Sпр | рН | 0.74 | 61 | 54.8 |
| (7озер*) | Хл. а | 0.63 | 30 | 21.1 |
| Hb | рН | 0.54 | 80 | 343.9 |
| (37 озер) | Площадь озера, км2 | 0.37 | 13 | 4.7 |
| Nобщ | ANC | 0.94 | 62 | 8.4 |
| (10 озер *) | Ptot | 0.75 | 31 | 7.6 |
Укрупнение организмов зоопланктона, вызванное преобладанием ракообразных (преимущественно кладоцер) над коловратками, представляет наиболее распространенный результат закисления водоемов (Schindler, 1994). Его часто отмечают в озерах, в которых вследствие ацидификации исчезли рыбы (Brett, 1989). В исследованных озерах при рН <5.3 обитал окунь. Тем не менее, отмечено увеличение средней индивидуальной массы (wср) и доминирование относительно крупных (l = 0.8-1.3 мм, w = 15-75 мкг) форм зоопланктона (рис. 3). В целом, в ацидных озерах доминирование ракообразных приводило к снижению Nобщ зоопланктона до <300 экз./л и росту wср до 20±0.5 мкг, в нейтральных зарегистрировано >1000 экз./л и 5±2 мкг соответственно. Для малых болотных озер wср = 42.2 – 15.6 lg(Nобщ).
Средняя за сезон и летняя биомасса (Вобщ) зоопланктона в большинстве случаев достоверно не различалась даже при сравнении водоемов с крайними значениями уровня рН воды. Распределение частоты встреч Вобщ в нейтральных и ацидных озерах также достоверно не различалось (χ2эмп 5.1> χ2станд 14.1, p < 0.05), в обоих случаях преобладали значения <2 г/м3 (55-56%) и 2-4 г/м3 (24-28%). Таким образом, в озерах Европейской России снижение численности зоопланктона при закислении фактически полностью компенсируется его укрупнением.
Трофические сети зоопланктона закисленных озер сильно упрощены (табл. 4). Небольшое число видов в них представлено в основном фильтраторами-фитофагами с крайне низкой плотностью межвидовых связей. По сравнению с нейтральным озером сокращается количество хищников. Из хищников присутствует только Polyphemus pediculus L. Крупные планктонные хищники (роды Leptodora, Bythotrephes, Cyclops, Heterocope) отсутствуют или редки в большинстве озер с рН <5-6 (Roff, Kwiatkowsky, 1977; Лазарева, 1994; Иванова, 1997; Вандыш, 2002). Сильно упрощенные трофические сети зоопланктона в пелагиали закисленных озер отмечают также для других регионов Европы и Северной Америки (Иванова, 1999; Havens, 1991; Locke, Sprules, 1994a). Считают, что это связано со снижением видового разнообразия при закислении (Havens, 1991; Locke, Sprules, 1994a; Лазарева и др., 2003а).
Таблица 4. Структура трофической сети зоопланктона, зообентоса и литоральных сообществ (по: Лазарева и др., 2003а,б).
| Параметры сети | рН<5 | pH>7 | |||||||
| Оз. Мотыкино | Оз. Дубровское | Оз. Хотавец | |||||||
| П | Б | Л | П | Б | Л | П | Б | Л | |
| Количество нехищных видов | 4 | 4 | 3 | 4 | 2 | 4 | 5 | 3 | 9 |
| Количество хищных видов | 1 | 11 | 15 | 2 | 8 | 16 | 4 | 5 | 8 |
| Количество хищников высшего порядка | 1 | 1 | 2 | 1 | 1 | 2 | 2 | 1 | 1 |
| Значение хищничества, % | 20 | 73 | 83 | 33 | 80 | 80 | 44 | 63 | 47 |
| Доля факультативных хищников, % | 0 | 47 | 22 | 17 | 63 | 20 | 22 | 38 | 12 |
| Доля каннибалов, % | 20 | 13 | 16 | 17 | 20 | 10 | 22 | 0 | 18 |
| Количество связей “хищник-жертва” | 4 | 88 | 67 | 6 | 40 | 82 | 19 | 16 | 50 |
| Плотность связей “хищник-жертва” | 0.8 | 5.9 | 3.7 | 1.0 | 4.0 | 4.1 | 2.0 | 2.0 | 2.9 |
| Модальное число трофических уровней | 3 | 6 | 6 | 4 | 5 | 5 | 4 | 4 | 7 |
| Генерализация | 5±0 | 81 | 41 | 42 | 51 | 41 | 4.81 | 31 | 72 |
| Количество генерализующих видов | 1 | 4 | 7 | 1 | 5 | 6 | 3 | 2 | 3 |
Примечание: П – планктон, Б – бентос, Л – литоральные сообщества.
При рН воды <5 более половины видов в бентосной сети и до 100% в литоральной составляют гетеротопные личинки насекомых. При рН >6, напротив, преобладают гомотопы (моллюски, пиявки, олигохеты, ракообразные). Распространение насекомых в закисленных водоемах вызвано снижением пресса рыб, благоприятным кислородным режимом у дна и конкурентным преимуществом гетеротопов перед гомотопами в экстремальных условиях обитания. Развитие в донных сообществах большого числа хищных видов во многом связано с обилием зоопланктона, который в толще воды слабо выедается хищниками. Эффективность использования энергии в длинных пищевых цепях ацидных водоемов повышается за счет распространения факультативных и генерализующих хищников (Лазарева и др., 2003б). Во всех озерах независимо от уровня закисления степень сложности структуры и архитектуры сети, длина пищевой цепи и генерализация возрастают в ряду зоопланктон → бентос → литоральные зооценозы (Лазарева и др., 2003б).
Механизм влияния закисления вод как комплексного процесса на зоопланктон малых озер Европейской России показан на рис. 4, темным выделены главные причинно-следственные взаимодействия. Экологические последствия природной (водами болот) и антропогенной ацидификации различны, поскольку различен сам механизм природного и антропогенного закисления (Комов, Лазарева, 1994; Лазарева и др., 1998, 2000а; Emmet et al., 1994).
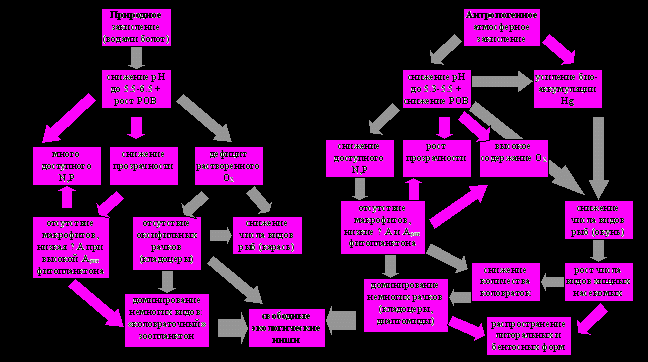
Рис. 4. Механизм влияния природного и антропогенного закисления на сообщества зоопланктона малых болотных озер.
Глава 5. Флуктуации и сукцессии зоопланктона, вызванные динамикой климата в регионе
Флуктуации климата включают периодичности от 2-3 лет до «вековых» циклов (~ 80 лет) и более (Шнитников, 1969; Адаменко, 1985). Выявление их влияния на динамику сообществ требует регулярных длительных и непрерывных рядов наблюдений, которые существуют для немногих водоемов. В бассейне Верхней Волги к таким относится Рыбинское водохранилище. Изменчивость зоопланктона водохранилища во времени и пространстве складывается из различий состава доминантных видов, общей структуры сообщества и уровня количественного развития (Nобщ, Вобщ). Ранее основное внимание уделяли особенностям пространственного распределения состава и обилия зоопланктона (Киселева, 1954; Мордухай-Болтовская, 1956; Воронина, 1959; Монаков, Семенова, 1966; Рыбинское водохранилище.., 1972; Ривьер, 1988, 1993, 2000), а также динамике Nобщ и Вобщ (Луферова, Монаков, 1966; Ривьер и др., 1982; Ривьер, 1988, 2000). Автором акцентировано внимание на закономерностях многолетней изменчивости состава, структуры зоопланктона и иерархии периодичностей колебаний характеристик сообщества (Лазарева, 1997, 2005а; Лазарева и др., 2001).
Полный список зоопланктона водохранилища включает >350 видов (Экологические проблемы…, 2001; 2007б). К широко распространенным (80-100%) проб относятся >25 видов (Лазарева, 2007а), 11 видов входят в состав доминантов Главного плеса (табл. 5). Состав и структура доминантного комплекса зоопланктона водохранилища сравнительно лабильны. Каждые 10-20 лет, в сообществе наблюдается частичное или полное замещение одного из главных доминантов или субдоминантов: Bosmina longispina ↔ B. coregoni, Conochilus hippocrepis ↔ C. unicornis, Eudiaptomus gracilis ↔ E. graciloides. Изменения такого рода с некоторым запаздыванием следуют за сменой фаз гидрологического цикла (Лазарева и др., 2001). Как правило, в них участвуют близко родственные таксоны: подвиды сменяют друг друга внутри вида, виды – внутри рода. В 1997-2004 гг. локально стали многочисленными 14 видов, которые в 80-х годах ХХ в. не входили в число доминантных, 5 прежних доминантов стали малочисленными (Лазарева, 2005а). С начала 90-х годов ХХ в. снизилась численность большинства видов коловраток, особенно видов родов Asplanchna, Brachionus, а также Keratella cochlearis Gosse (Лазарева и др., 2001; Лазарева, 2005а,в, 2007а).
Таблица 5. Доминантные виды (июнь-август) зоопланктона центральной части Рыбинского водохранилища («стандартные» станции) в 2003-2005 гг.
| Ракообразные | %Ncrust | Коловратки | %Nrot |
| Mesocyclops leuckarti (ad.+juv.) | 46±5 | Conochilus hippocrepis | 228 |
| Daphnia galeata | 122 | Polyarthra major | 206 |
| Eudiaptomus gracilis | 92 | Synchaeta pectinata | 156 |
| Thermocyclops oithonoides (ad.+juv.) | 8±1 | Kellicottia longispina | 92 |
| Bosmina longispina | 53 | Keratella cochlearis | 92 |
| | | Keratella quadrata | 84 |
В течение 1956-2005 гг. для структуры и обилия зоопланктона Рыбинского водохранилища выявлены периодичности трех порядков. Циклы продолжительностью 6-7 лет характерны для динамики видового богатства (Sп), 10-11 лет – для Nобщ, Вобщ и характеристик таксономической структуры сообщества (индексы ISimp, Hb, Е), медленные колебания (~20 лет) отмечены для Вобщ и wср (рис. 5), вероятно, такие же свойственны видовой плотности (Sп) (Лазарева, 1997, 2007а; Лазарева и др., 2001).
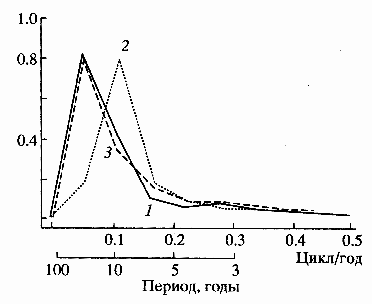
Рис. 5. Нормированные периодограммы колебаний Вобщ (1), Nобщ (2) и wср (3) в Рыбинском водохранилище (по: Лазарева и др., 2001).
Наибольший уровень доминирования и, соответственно, наименьший уровень разнообразия в сообществе соответствует максимумам солнечной активности по числам Вольфа (Лазарева, 1997). Максимальные значения трофического индекса Е зоопланктона следуют за таковыми концентрации хлорофилла а с запаздыванием на 2 года (Лазарева, 1997). Колебания концентрации пигмента происходят фактически синхронно с динамикой поступления суммарной солнечной радиации (Пырина, 2000). Напротив, периодичность изменения видового богатства зоопланктона близка к характерной для стока рек бассейна Верхней Волги, наиболее отчетливо прослеживаются колебания с периодами 6-7 и 8-12 лет (Лазарева, 1997).
Пики Nобщ зоопланктона отмечены в годы с максимальной температурой воды (рис. 6). До 1985 г. они совпадали с подъемами содержания хлорофилла а, после зарегистрировано их расхождение. Колебания в противофазе показателей фито- и зоопланктона известны также для оз. Красного (Трифонова, Макарцева, 2006). В Рыбинском водохранилище максимум Вобщ зоопланктона пришелся на многоводную фазу гидрологического цикла (1977-1995 гг.), в которой объем притока вод был наибольшим за вековой период наблюдений (Литвинов, Рощупко, 2000). До середины 70-х годов прошлого века увеличение Вобщ зоопланктона происходило на фоне снижения притока воды, в последующие годы – следовало за изменением притока (рис. 7). Синхронизация колебаний Вобщ и притока в 20-летнем цикле произошла, вероятно, под влиянием биостока из Шекснинского водохранилища, заполненного в 1963 г. В настоящее время он заметно влияет на зоопланктон в Шекснинском и Главном плесах Рыбинского водохранилища (Ривьер, 1998; Лазарева, 2005а). В целом, в маловодный период 1963-1976 гг. Вобщ зоопланктона была в 2.4-2.7 раза ниже, численность рачков – в 1.8-1.9 раза ниже по сравнению с многоводным периодом 1977-1990 гг. (Лазарева и др., 2001). Однако в 10-летнем цикле пики биомассы в 1963-1964, 1971-1973, 1980-1982, 1987-1988, 1994-1995 и 2005 гг. в основном совпадали с маловодными периодами (рис. 7).
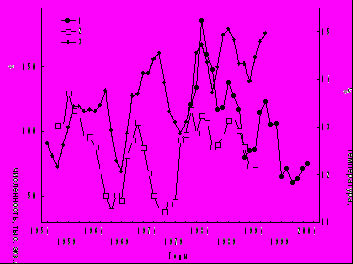
Рис. 6. Динамика Nобщ зоопланктона и температуры воды в Рыбинском водохрани-лище в 1954-2005 гг. (сглаженные кривые при 3-летнем усреднении).
1 – численность зоопланктона в августе, 2 – то же среднее май-октябрь, 3 – температура воды (данные Гидрометеослужбы).
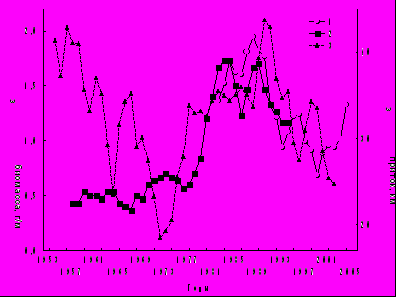
Рис. 7. Динамика Вобщ зоопланктона и годового притока воды в Рыбинском водохранили-ще в 1953-2005 гг. (сглаженные кривые при 3-летнем усреднении).
1 – Вобщ зоопланктона в августе, 2 – то же среднее май-октябрь, 3 – годовой приток воды (по: Литвинов, Рощупко, 2007).
Динамика хлорофилла, отражающая межгодовые колебания доступных для зоопланктона трофических ресурсов, определяет >60% вариаций Вобщ зоопланктона, вклад изменений термического и гидрологического режимов водохранилища составляет 4-7% (табл. 6). Представленная регрессионная модель объясняет >80% вариаций Вобщ и может быть использована для приблизительной оценки количества зоопланктона в водоеме: Вобщ = 0.07 Хл. + 0.05 Приток + 0.13 Тводы – 0.47 ά – 2.03
Таблица 6. Факторы, определяющие многолетнюю динамику биомассы зоопланктона (Вобщ) Рыбинского водохранилища (n = 42).
| Фактор | R | F | R2,% |
| Хл. | 0.72 | 46.9 | 66 |
| Приток | 0.56 | 5.7 | 7 |
| ά | –0.41 | 4.2 | 4 |
| Тводы | 0.43 | 4.0 | 4 |
Примечание: n – количество наблюдений (проб), Хл. – концентрация хлорофилла а (по: Пырина, 2000; Минеева, 2006), ά – коэффициент условного водообмена. Остальные обозначения как в табл. 3.
На фоне квази-циклических колебаний отмечены направленные изменения численности ряда видов и таксонов более высокого ранга зоопланктона водохранилища. Изменение относительного обилия доминантов наиболее выражено при сравнении маловодных периодов (табл. 7), а численности и биомассы крупных таксономических групп и сообщества в целом – при сравнении многоводных (табл. 8). Для некоторых показателей (Nrot, Ncr/Nrot) выявлены достоверные различия по маловодным и многоводным периодам. В динамике зоопланктона доминируют изменения, отражающие влияние внешних для экосистемы факторов: тренды по многоводным периодам выражены сильнее, чем по маловодным (табл. 7 и 8). Отмеченные изменения в зоопланктоне свидетельствуют в основном о деэвтрофировании экосистемы.
Таблица 7. Динамика численности некоторых реперных видов зоопланктона Рыбинского водохранилища в 1956-2004 гг.
| Вид | Относительная численность,% | Тренд | ||||
| | маловодные | многоводные | маловодные | (+/–), | ||
| | 1961-1965 | 1971-1975 | 1981-1985 | 1991-1995 | 2003-2005 | % |
| Keratella quadrata | 9 | 14 | 15 | 14 | 8 | нет |
| Kellicottia longispina | 11 | <5 | <5 | 5 | 9 | нет |
| Thermocyclops oithonoides | 7 | 16 | 6 | 17 | 8 | нет |
| Bosmina longispina | 11 | <5 | 17 | 17 | 5 | нет |
| Eudiaptomus sp. | <5 | 7 | 5 | 5 | 9 | +30 |
| Daphnia galeata | <5 | <5 | <5 | 5 | 12 | +200 |
| Polyarthra major | <1 | <1 | <5 | 8 | 20 | +2000 |
| Polyarthra vulgaris | 12 | <5 | 7 | <5 | <5 | –200 |
| Chydorus sphaericus | 7 | 11 | 6 | 6 | <5 | –75 |
| Sобщ | 164 | – | – | – | 364 | +120 |
Примечание: Sобщ – общее число видов в фаунистическом списке: исходное (по: Рыбинское водохранилище, 1972), конечное (по: Лазарева, 2007б).
Снижение почти вдвое численности коловраток, скорее всего, представляет следствие их конкуренции с дрейссенами за ресурсы сестона (Лазарева, 2005б; 2007а). В присутствии дрейссены потери взвеси от общего ее содержания в толще воды увеличиваются в 8 раз (Деренговская и др., 2002). Заметное влияние другого вселенца планктофага тюльки на структуру и количество зоопланктона выявлено только в местах скопления рыб в период их интенсивного питания в июне (Кияшко и др., 2007). Низкая численность доминирующего планктофага в водоеме представляет одну из основных причин массового развития в планктоне крупных рачков (роды Daphnia, Leptodora, Bythotrephes).
Таблица 8. Направленные изменения общих характеристик зоопланктона в Рыбинском водохранилище в 1956-2004 гг.
| Показатель | Многоводные периоды (1, n = 94)* | Маловодные периоды (2, n = 38)** | Тренд (+/–),% | |||
| | 1956-1962 | 1989-1995 | 1963-1969 | 1996-2004 | 1 | 2 |
| Вобщ, г/м2 | 2.9±0.2 | 6.9±0.4 | – | – | +140 | – |
| Вобщ, г/м3 | 0.46±0.04 | 1.31±0.08 | 0.79±0.09 | 0.87±0.12 | +185 | +10 |
| Nclad | 11.2±0.8 | 16.3±1.4 | 10.4±2.3 | 9.9±1.6 | +45 | –5 |
| Ncop | 19.4±1.7 | 31.7±4.1 | 20.2±1.7 | 26.9±4.4 | +60 | +30 |
| Nrot | 79.4±10.4 | 41.8±5.1 | 76.7±7.3 | 26.2±8.6 | –90 | –90 |
| Ncr/ Nrot | 0.4 | 1.1 | 0.4 | 1.4 | +175 | +250 |
| Bz/Bph (V-X) | 0.34±0.04 | 0.74±0.07*** | 0.34±0.03 | 0.41±0.03 | +120 | +20 |
Примечание: * – среднее V-X в конце каждого мно-говодного периода. ** – среднее VI («пик») в начале каждого маловодного периода.*** – данные 1981-1986 гг.
Помимо трендов количественных характеристик выявлены качественные сдвиги в структуре планктона водохранилища: расширение фаунистического списка видов, смены доминантов, например: Polyarthra vulgaris → P. major, Bosmina longispina → Daphnia galeata (табл. 7) и групп планктона Rotatoria → Crustacea (табл. 8), а также вселение и распространение новых для водоема видов. В 2001-2004 гг. в Рыбинском водохранилище отмечено 8 новых видовзоопланктона (Лазарева, 2005а,в). Интенсивно распространяются в бассейне Верхней Волги Diaphanosoma orghidani Negrea и Asplanchna henrietta Langhaus (Лазарева, 2005а, 2008). Оба вида южного происхождения, ранее отмечены в водоемах средней и нижней Волги (Коровчинский, 1986; Тимохина, 2000). В современный маловодный период (с 1996 г.) отмечено увеличение численности >10 видов, новых и впервые зарегистрированных в водоеме в 70-80-х годах прошлого века (Лазарева, 2005а,в). Некоторые из них стали многочисленными в течение 2-3 лет в водохранилищах и озерах бассейна, в сукцессионном масштабе фактически одновременно. Это подтверждает ведущее значение региональных эдафических, в частности, климатических факторов для динамики обилия новых и редких планктонных форм.
Особенностью натурализации видов в водохранилищах Верхней Волги служит первоначальное расселение их в прибрежье и устьевых участках рек (Лазарева, 2008), тогда как южнее виды-вселенцы встречаются почти исключительно в пелагиали за пределами зоны зарослей и речных эстуариев (Попов, 2005; Романова и др., 2005). Натурализация включает несколько этапов (Лазарева, 2008): 1 – единичные находки вида как следствие случайного заноса; 2 – формирование локальных скоплений в литорали, заливах, устьях рек-притоков; 3 – увеличение численности вида в открытом водохранилище; 4 –локальное, реже полное по всей акватории, вытеснение аборигенных близкородственных видов из доминантного комплекса зоопланктона.
Для зоопланктона двух озер, ацидного (рН <5) мезотрофного оз. Дубровское и нейтрального (рН >7) эвтрофного оз. Хотавец (бассейн Рыбинского водохранилища), за 13 лет наблюдений удалось выделить лишь циклические сукцессии, связанные с динамикой климатических и гидрофизических характеристик. Периодичность колебаний гидрофизических характеристик, а также Nобщ и Bобщ зоопланктона в озерах близка к 10-летнему циклу. Этот цикл четко выражен в динамике абиотических и биотических характеристик Рыбинского водохранилища (Литвинов и др., 2005; Пырина, 2000; Лазарева, 1997; Лазарева и др., 2001). Отражением медленных смен состава зоопланктона служит появление в 2002 г. в оз. Хотавец новой коловратки Asplanchna henrietta, почти одновременно с ростом ее численности в Рыбинском водохранилище.
Хотя исследованные озера принимают бедный в фаунистическом отношении диффузный сток из болот, выявлена тенденция к увеличению большинства характеристик видового богатства зоопланктона в многоводный период, что характерно также для Рыбинского водохранилища (Лазарева, 1997). В сборах планктона на пике водности (1987-1992 гг.) достоверно возрастает количество случайных видов (F = 5.9, p < 0.005), что, вероятно, связано с усилением выноса в центральную часть озер организмов из литорали, зарослей и донных биотопов при увеличении проточности в многоводные годы. Зарегистрировано также изменение состава и структуры доминантных комплексов зоопланктона, изменение встречаемости и численности сукцессионных видов, снижение фаунистического индекса трофности Е и показателей динамичности фауны (Lazareva, 2000). Как и в Рыбинском водохранилище, наибольшие изменения состава и структуры доминантного комплекса зоопланктона озера выявлены при переходе от многоводного периода гидрологического цикла к маловодному.
В оз. Хотавец наибольшую летнюю Nобщ регистрировали на пике и спаде водности водоемов бассейна Верхней Волги (1987-1996 гг.), в то время как уровень Bобщ в сравниваемые периоды достоверно не различался. Напротив, в оз. Дубровское отмечена тенденция к увеличению обилия зоопланктона в маловодный период (1998, 2002 гг.) и на подъеме водности (1984-1986 гг.). Различный ход многолетней динамики зоопланктона в озерах разных типов, вероятно, связан с несходством направленности изменения уровня рН воды. Максимум рН в нейтральном водоеме соответствовал минимуму этого показателя в закисленном. Пики Nобщ и Bобщ зоопланктона в оз. Хотавец запаздывали на 3-4 года относительно таковых в сообществе Рыбинского водохранилища, Nобщ положительно коррелировала с динамикой температуры воды (r = 0.70, p <0.05). Запаздывание отклика экосистем болотных водоемов на изменение общей увлажненности в регионе, вероятно, связано гидрологической инертностью болотных водосборов, для которых отмечают более высокий (на 10-15%) по сравнению с лесными уровень эвапотранспирации и большую влагоемкостью торфяников (Иванов, 1957; Богословский и др., 1984).