
Биотехнология методы получения аминокислот и белков, меченых стабильными изотопами
| Вид материала | Документы |
- Высокомолекулярные азотосодержащие органические вещества, молекулы которых построены, 51.51kb.
- Разработка методов биотехнологического получения белков, аминокислот и нуклеозидов,, 735.56kb.
- O в молекулы аминокислот и белков. О. В. Мосин Московская государственная академия, 401.15kb.
- Календарно-тематический план лекций по биологической химии для студентов II курса медико-профилактического, 39.13kb.
- В строении белков одно общее: они состоят из аминокислот. Всего в состав молекул, 1313.07kb.
- Свойства белков и аминокислот, 100.39kb.
- Первый. Белки под большой молекулярной массой от нескольких десятков до нескольких, 28.19kb.
- Влаконичной форме дайте письменные ответы на следующие вопросы: Значение белков, аминокислот, 23.55kb.
- Быстроусваиваемый полный комплекс аминокислот, полученный из сывороточных белков. Предназначен, 177.83kb.
- Биотехнология масс-спектрометрическая оценка уровня включения дейтерия и углерода-13, 335.24kb.
1 2
Табл. 1
Параметры роста некоторых используемых в биотехнологии штаммов метилотрофных бактерий
| Штаммы бактерий | Молярный выход биомассы, г/моль метанола | Удельная скорость роста, ч-1 | Эффективность конверсии углерода метанола, % | Количество потребленного азота, % |
| Рибулозо-5-монофосфатный путь ассимиляции углерода | ||||
| Pseudomonas C1 | 17,3 | 0,49 | 67,5 | 13,2 |
| Pseudomonas methanolica | 17,0 | 0,63 | 66,5 | 11,0 |
| Methylomonas methanolica | 15,7 | 0,52 | 62,0 | 11,7 |
| Сериновый путь ассимиляции углерода | ||||
| Pseudomonas 1 | 12,1 | 0,176 | 47,5 | 11,37 |
| Pseudomonas 135 | 12,1 | 0,14 | 47,5 | 11,37 |
| Pseudomonas AM1 | 9,8 | 0,093 | 37,6 | 11,20 |
| Pseudomonas M27 | 13,1 | 0,108 | 51,0 | 9,40 |
| Pseudomonas roseus | 13,1 | 0,15 | 51,0 | 10,60 |
Другими перспективными источниками изотопномеченых аминокислот и белков признаны метилотрофные микроорганизмы, которые в таксономическом аспекте представлены грамположительными, грамотрицательными бактериями и дрожжами, интерес к которым в настоящее время все возрастает благодаря разработке новых технологий химического синтеза метанола [102]. Эти бактерии привлекают внимание исследователей прежде всего как дешевые источники микробного белка и аминокислот [103, 104]. Знание путей бактериального метаболизма позволяет осуществлять направленное введение изотопной метки в молекулы аминокислот. Как известно, метилотрофные бактерии окисляют метанол с использованием фермента - метанолдегидрогеназы, последующие окислительные реакции катализируют формальдегид- и формиатдегидрогеназа [105-108]. Лишь затем продукт окисления метанола в виде формальдегида фиксируется клеткой одним из двух путей ассимиляции углерода: рибулозо-5-монофосфатным и сериновым [109, 110]. Основные параметры роста некоторых используемых в биотехнологии штаммов метилотрофных бактерий представлены в таблице 1.
Ауксотрофные штаммы метилотрофных бактерий начали эффективно использовать для получения [2H]- и [13C]аминокислот. Для этих целей перспективно осуществлять биологическую конверсию дешёвых низкомолекулярных меченых субстратов - (13С)метанола, (2Н)метанола и 2Н2O в дорогостоящие меченые БАС в клетках метилотрофов [111-113]. Традиционным подходом при этом остаётся выращивание соответствующих штаммов-продуцентов аминокислот, устойчивых к росту на средах, содержащих стабильные изотопы водорода, углерода, азота и др. В работах [114, 115] сообщается о получении [13C]аминокислот (уровни включения стабильных изотопов в молекулах варьируют от 30% для L-[13C]лейцина до 90% для L-[13C]фенилаланина) за счёт использования ауксотрофных по L-изолейцину бактерий Methylobacillus flagellatum.
Заслуживает упоминания то, что (13С)метанол в отличие от 2Н2О не оказывает существенного биостатического эффекта на ростовые и биосинтетические характеристики метилотрофов [116], поэтому данный подход можно эффективно использовать для введения в молекулы синтезируемых БАС двойной изотопной метки (например, введение изотопа 13С в молекулы на фоне максимальных концентраций 2Н2О в ростовых средах). В работе [117] были получены [2H]- и [13С]аминокислоты с разными уровнями изотопной обогащённости при росте ауксотрофного по L-лейцину штамма факультативных метилотрофных бактерий Brevibacterium methylicum и ауксотрофного по L-изолейцину штамма облигатных метилотрофных бактерий Methylobacillus flagellatum на минимальных средах с (13С)метанолом, (2Н)метанолом и 2Н2О. [13C]- и [2Н]аминокислоты разного уровня изотопной замещённости выделяли как из культуральных жидкостей, полученных после выращивания бактерий на средах с соответствующими изотопномечеными субстратами, так из гидролизатов белков биомассы. Биосинтетически полученные [2H]- и [13С]аминокислоты представляли собой смеси, в которых присутствовали изотопнозамещённые формы молекул, различающиеся количеством атомов водорода и углерода, замещённых на 2Н и 13С. При этом распределение зависело как от общего включения изотопа в молекулу, так и от способа их получения. Данные по суммарным уровням включения стабильных изотопов в молекулы секретируемых аминокислот и аминокислотные остатки суммарных белков биомассы B. methylicum и M. flagellatum представлены в таблице 2. Эти исследования показали, что в условиях ауксотрофности по лейцину уровень изотопного обогащения лейцина, а также метаболически связанных с ним аминокислот немного ниже, чем для других аминокислот, вероятно, за счёт сохранения минорных путей метаболизма, связанных с биосинтезом данных аминокислот de novo. Так, при выращивании B. methylicum на среде, содержащей 98% 2Н2О и немеченый L-лейцин, уровни включения дейтерия в индивидуальные аминокислоты культуральной жидкости составили 51% для лейцина/изолейцина, 58,8% для валина, в то время как уровни изотопного включения для аланина составили 77,5%, для фенилаланина -75% (табл. 2).
Табл. 2
Суммарные уровни включения стабильных изотопов в молекулы секретируемых аминокислот и аминокислотные остатки суммарных белков биомассы B. methylicum* и M. flagellatum**.
| А      минокислоты минокислоты | Концентрация 2Н2О в ростовой среде, об% 24,5 49,0 73,5 98,0 КЖ# белок КЖ белок КЖ белок КЖ белок | 1 %13СН3ОН КЖ белок | |||
| Глицин | - 15,0 | - 35,0 | - 50,0 | - 90,0 | 60,0 90,0 |
| Aланин | 24,0 20,0 | 37,5 45,0 | 62,5 62,5 | 77,5 97,5 | 35,0 95,0 |
| Валин | 20,0 15,0 | 46,3 36,3 | 43,8 50,0 | 58,8 50,0 | 50,0 50,0 |
| Лейцин/изолейцин | 15,0 10,0 | 47,0 42,0 | 46,0 45,0 | 51,0 49,0 | 38,0 49,0 |
| фенилаланин | 15,0 24,5 | 27,5 37,5 | 51,2 50,0 | 75,0 95,0 | 95,0 80,5 |
| Tирозин | - 20,0 | - 25,6 | - 68,8 | - 92,8 | - 53,5 |
| Серин | - 15,0 | - 36,7 | - 47,6 | - 86,6 | - 73,3 |
| Aспарагиновая кислота | - 20,0 | - 36,7 | - 60,0 | - 66,6 | - 33,3 |
| Глутаминовая кислота | - 20,0 | - 40,0 | - 53,4 | - 70,0 | - 40,0 |
| Лизин | - 10,0 | - 35,3 | - 40,0 | - 58,9 | - 54,4 |
*Данные по включению дейтерия в молекулы аминокислот приведены для B. methylicum при росте на средах, содержащих 2% CH3OH и 24,5; 49,0; 73,5; 98,0% 2Н2О
**Данные по включению 13С приведены для M. flagellatum при росте на среде, содержащей обычную воду и 1% 13СН3ОН.
#Термином КЖ обозначены культуральные жидкости, полученные после отделения клеток из ростовых сред
Аналогичная корреляция наблюдалась и в аминокислотах белковых гидролизатов. Уровни включения 2Н и 13С в метаболически связанных аминокислотах в пределах одинаковых концентраций меченых субстратов, обнаружили определённую коррелляцию: уровни изотопного включения для валина и лейцина (семейство пирувата), фенилаланина и тирозина (семейство ароматических аминокислот) коррелировали (табл. 2). Уровни изотопного включения для глицина и серина (семейство серина), аспарагиновой кислоты и лизина (семейство аспарагина) также имели близкие величины. Важным результатом являются высокие уровни включения стабильных изотопов 2Н и 13С в молекулы полученных аминокислот. В настоящее время исследования по изучению биотехнологического потенциала метилотрофных бактерий для направленного синтеза изотопномеченых аминокислот и других БАС продолжаются.
Генно-инженерные методы получения изотопномеченных аминокислот и белков.
Осуществлять направленное биосинтетическое получение индивидуальных белков, меченных стабильными изотопами удобно за счёт использования векторов экспрессии нужных генов, ответственных за биосинтез того или иного интересующего исследователей белка. Оправдано и целесообразно использование для этих целей векторов экспрессии на основе плазмидной ДНК бактерии E. coli, например, вектор экспрессии Т4 лизоцима, включающий в своем составе плазмиду pHSe5 [118]. В результате использования этого вектора экспрессии, были получены миллиграммовые количества Т4-лизоцима, селективно меченного стабильными изотопами азота 15N или углерода 13C. Включение стабильных изотопов в данном случае удалось осуществить за счет роста генного конструкта E. coli на средах, содержащих [15N]- или [13С]аминокислоты. Метод также может применяться для получения индивидуальных меченых белков, экспрессия которых происходит в системах, отличных от E. coli, например, системы экспрессии на основе клеток насекомых или млекопитающих [119].
Другие микробные системы, в которых белки экспрессируются с высокими выходами, также могут быть пригодны для получения изотопномеченых аналогов белков. К ним относятся, прежде всего такие хорошо изученные биологические объекты, как дрожжи, бактерии и бактериофаги. Так, за счёт использования вышеперечисленных микробных объектов в качестве векторов экспрессии были получены препаративные количества индивидуальных очищенных [15N]белков: нуклеаза стафилококка [120], интерлейкин 1 [121], белок репрессор фага P22C2 [122], тиредоксин E. coli [123], гемоглобин [124], -протеаза [125], ингибитор субтилизина [126], репрессор фага [127], и белок человеческого фактора роста N-ras P21 [128].
В работе [129] описан метод получения индивидуальных [2H]белков с использованием вектора экспрессии на основе штамма облигатных метилотрофных бактерий Methylobacillus flagellatum. Метод состоит в том, что в метилотрофах клонируют структурный ген исследуемого белка. Таким методом можно получать, например, [2H]-интерфероны, хорошо экспрессируемые в клетках метилотрофов, либо другие интересующие исследователей белки. Метод также позволяет вводить в молекулы аминокислот и белков другие стабильные изотопы, например, 13С. В связи с этим следует подчеркнуть, что основным недостатком при использовании полученных данным методом [13C]аминокислот в ЯМР-исследованиях являются всё же недостаточно высокие уровни изотопного обогащения аминокислот, что обусловливает усложнение спектров ЯМР за счет 12C- 13C-спин-спинового взаимодействия между близлежащими атомами углерода в молекуле [130]. Так как мультиквантовые резонансы близлежащих атомов углерода в молекуле являются основным препятствием для интерпретации спектров ЯМР, необходимо применять усовершенствованные методы получения [13C]аминокислот, позволяющие лимитировать процесс разбавления метки. Так, в последнее время были генетически сконструированы новые штаммы бактерий, которые несут мутации по генам метаболизма определенного круга предшественников этих аминокислот [131]. Это позволяет избежать разбавления метки при росте микроорганизма на среде, содержащей те или иные меченые субстраты за счет ингибирования биосинтеза аминокислот de novo у данных мутантных штаммов бактерий.
При выборе определенных мутаций по генам метаболизма стремятся удовлетворить как миниум двум условиям для нормального функционирования подобных генетически сконструированных систем, чтобы, во-первых, по возможности снизить деградацию метки или ее разбавление в процессе внутриклеточного синтеза немеченых предшественников de novo и во-вторых, свести к минимуму процессы перестройки меченых положений углеродного скелета молекулы за счет биосинтеза одинаковых интермедиантов, образующихся по сопряжённым путям биосинтеза. Данная стратегия реализована в работе [132], где сообщается о получении двух генетически сконструированных штаммов бактерий, обозначенных как E. coli DL10 и E. coli DL11, которые несли геномные делеции, исключающие обмен атомов углерода между интермедиаторами в процессе гликолиза и в цикле трикарбоновых кислот.
За счёт использования данных штаммов удалось получить препаративные количества [13C]аминокислот с уровнями изотопного обогащения до 95%. Схема, иллюстрирующая мутации в генах метаболизма в цикле гликолиза и трикарбоновых кислот, приведена на рис. 2. Оба сконструированных штамма бактерий E. coli имели в геноме мутации, затрагивающие гены семи ферментов основных путей метаболизма (рис. 2).
Ферменты (1-5) у штамма E. coli DL10 были инактивированы за счёт мутаций, вследствие чего он ассимилировал в качестве источников углерода и энергии сукцинат и ацетат из ростовой среды, а [1-13C]лактат добавляли в ростовую среду для компенсации метаболических потребностей клетки и для введения 13C-метки в молекулы аминокислот, синтезируемых в процессе гликолиза.
Другой штамм бактерий E. coli DL11 мог утилизировать немеченую глюкозу в качестве источников углерода и энергии по гликолитическому пути ассимиляции углерода, в то время как [1,4-13C]cукцинат и [1-13C]ацетат добавляли в ростовую среду для того, чтобы стимулировать биосинтез [13C]аминокислот, образующихся по циклу трикарбоновых кислот. Кроме того, в этом случае было необходимо ввести в бактериальный геном дополнительную мутацию, связанную c геном -кетоглутаратдегидрогеназы, чтобы минимизировать процесс деградации метки в цикле трикарбоновых кислот.
6
 -фосфоглюконовая кислота
-фосфоглюконовая кислота


 рибулозо-5-фосфат глюкоза
рибулозо-5-фосфат глюкоза  рибозо-5-фосфат глюкозо-6-фосфат
рибозо-5-фосфат глюкозо-6-фосфат 



 ксилулозо-5-фосфат фруктозо-6-фосфат
ксилулозо-5-фосфат фруктозо-6-фосфат
 седогептулозо-7-фосфат фруктозо-1,6-дифосфат
седогептулозо-7-фосфат фруктозо-1,6-дифосфат 
 эритрозо-4-фосфат фосфоглицеральдегид-3-фосфат 6 дегидроксиацетонфосфат
эритрозо-4-фосфат фосфоглицеральдегид-3-фосфат 6 дегидроксиацетонфосфат 

 фосфоенолпируват
фосфоенолпируват 





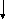 пируват лактат
пируват лактат 2 1
ацетил-СоА 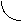 4 3
4 3 

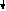 5
5 оксалоацетат цитрат
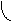
малат изоцитрат
фумарат СО2 -кетоглутарат
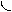 сукцинат 7
сукцинат 7 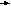 СО2
СО2 сукцинил-СоА
Рис. 2. Мутации в генах метаболизма цикла гликолиза и трикарбоновых кислот (по ЛеМастеру Д. М., 1982).
Обозначения ферментов: 1 - пируватдегидрогеназа; 2 - фосфоенолпируваткарбоксилаза; 3 - фосфоенолпируваткарбоксикиназа; 4 - NAD-малатдегидрогеназа; 5 - NADP-малатдегидрогеназа; 6 - триозофосфатизомераза; 7 - -кетоглутаратдегидрогеназа.
Выделение изотопномеченых аминокислот из белковых гидролизатов микроорганизмов.
Биомасса микроорганизмов, выращенных на средах, содержащих стабильные изотопы, является ценным источником различных изотопномеченых БАС, в том числе аминокислот. При этом наиболее распространённым и традиционным методом препаративного выделения аминокислот из клеточной биомассы является её гидролиз с использованием ферментативных или химических методов и последующая ионообменная хроматография на катионо- и анионообменных смолах (дауэкс, амберлит, пермутит, аминекс, дуолит и др.) [133]. Большое значение при проведении гидролиза белка имеет выбор того или иного гидролизирующего агента, который определяется целью исследования. Ферментативное расщепление протеолитическими ферментами может протекать ступенчато с концов молекулы (экзопептидазами) или путём расщепления специфических отдельных пептидных связей полипептидной цепи (эндопептидазами), причём специфичность зависит от конфигурации, аминокислотной последовательности и конформации белка [134]. Для селективного химического расщепления белков разработано очень много методов [135], среди которых имеется несколько методов расщепления по -углеродному атому (например, через остатки дегидроаланина). Щёлочи и кислоты обладают высокой гидролизующей способностью и поэтому их использование приводит к разрушению некоторых аминокислот и к изотопному обмену в триптофане, тирозине и гистидине и в некоторых других аминокислотах. В условиях щелочного гидролиза (4 н. Ba(OH)2 или NaOH, 24 ч, 1100) реакций изотопного обмена водорода на дейтерий практически не наблюдается (исключением является протон (дейтерон) у атома С2’ гистидина) [136]. Существенным недостатком щелочного гидролиза, лимитирующим его использование, является значительная рацемизация аминокислот. Поэтому для препаративных целей щелочной гидролиз используется крайне редко, в то время как кислотный - очень широко. Кислотный гидролиз в стандартных условиях (6 н. НCl или 8 н. Н2SO4, 24 ч, 1100), как известно, приводит к полному разрушению триптофана и частичному разрушению серина, треонина и некоторых других аминокислот [137]. Добавление в реакционную среду фенола [138], тиогликолевой кислоты [139], -меркаптоэтанола [140], позволяет сохранить до 80-85% триптофана. Кроме этого, в условиях кислотного гидролиза с высокой скоростью протекает изотопный обмен ароматических протонов (дейтеронов) в молекулах триптофана, тирозина и гистидина [141], а также протонов (дейтеронов) при атоме С3 аспарагиновой и С4 глутаминовой кислот [142]. Поэтому для получения реальных данных о биосинтетическом включении дейтерия в белок рекомендуется проводить кислотный гидролиз в присутствии дейтерированных реагентов. Этим способом могут быть выделены и анализированы с использованием ионообменной хроматографии большинство аминокислот в составе гидролизатов белка (рис. 3). Так, при помощи ионообменной хроматографии были препаративно выделены [2H], [13C]- и [15N]аминокислоты из белковых гидролизатов разных природных источников с выходами индивидуальных аминокислот от 77% до 95% и с уровнями изотопного включения, превышающими 95% [143]. Однако, метод выделения аминокислот из гидролизатов биомассы, будучи широко применяем на практике часто требует использования вредных буферных растворов (ацетат, формиат, пиридин и др.), нескольких колонок с последующей рехроматографией для полного выделения чистых аминокислот из гидролизатов биомассы.
Условия ионообменного выделения [2H]аминокислот из гидролизатов суммарных белков биомассы микроводоросли Scenedesmus obliquus, состав элюирующих растворителей, время проведения хроматографического анализа и др, были исследованы в работе [144]. При этом, уровни изотопного включения дейтерия в аминокислоты, выделенные из гидролизатов белков Scenedesmus obliquus составили более 98%. Вследствие протекания реакций обратного изотопного (1Н-2Н)-обмена с протонированным растворителем в ходе элюирования [2H]аминокислот с сорбента, протоны в -положении аспарагиновой кислоты и -положении глутаминовой кислоты были обогащены дейтерием на 90%, т. е. ниже, чем для других аминокислот. Подвижные атомы дейтерия в -положении имидазольного кольца молекулы гистидина и атомы дейтерия при гетероатоме азота в индольном кольце триптофана также легко обменивались на протоны в составе водных растворителей при выделении аминокислот.
[13C]аминокислоты были выделены из гидролизатов суммарных белков биомассы штамма метаногенных бактерий Methanobacterium espanolae при росте бактерий на [1-13C]- и [2-13C]ацетате с уровнями включения 13С в молекулы аминокислот до 90% [145]. Согласно цитируемым там данным, менее 2% случайной метки в молекулах аминокислот были распределены между атомами углерода в позициях, происходящих из 13С карбоксильной или метильной группы ацетата и еще меньший процент включения метки детектировался в положениях углеродного скелета молекул, образованных из 13СО2.
Большой практический интерес представляет реализация преимуществ препаративной обращенно-фазовой высокоэфективной жидкостной хроматографии (ОФ ВЭЖХ) при получении оптически чистых меченых аминокислот и их N-производных в количествах, необходимых для биоаналитических и синтетических целей [146, 147]. Так, в работе [147] описан метод препаративного выделения индивидуальных аминокислот из различных микробиологических источников с помощью ОФ ВЭЖХ в виде бензилоксикарбонильных производных (N-Cbz производных) аминокислот. Разработанный метод позволяет выделять аминокислоты с высоким выходом (от 67% до 89%) и хроматографической чистотой (96-99%) [147] и может быть использован для выделения [2H]-, [13C]-, [15N] и [18O]аминокислот из белковых гидролизатов различных источников.
ХИМИКО-ФЕРМЕНТАТИВНЫЙ МЕТОД ПОЛУЧЕНИЯ ИЗОТОПНОМЕЧЕНЫХ АМИНОКИСЛОТ И БЕЛКОВ.
Другим подходом по получению изотопномеченых аминокислот является химико-ферментативный метод, основанный на комбинации синтетических и ферментативных реакций. Для этого перспективно и экономически оправдано использование препаратов очищенных ферментов и их экстрактов, безклеточных ферментативных систем, а также иммобилизованных ферментов. Ферментативные реакции осуществляют на иммобилизованных ферментах, например, таких как аланиндегидрогеназе (КФ 1.4.1.1) в присутствии NADH при получении [2H]аланина [148], иммобилизованной на сахарозе фенилаланинаммонийлиазе (КФ 4.3.1.5) и фенилаланингидроксилазе (КФ 1.14.16.1), при получении [2H]фенилаланина [149] и [2H]тирозина [150], триптофансинтазе (КФ 4.2.1.20), при получении [2H]триптофана [151], глутаматдегидрогеназе (КФ 1.4.1.2), при получении [2H]глутаминовой кислоты [152], аспартазы (КФ 4.3.1.1), при получении [2Н]аспарагиновой кислоты [153] и серингидроксиметилазе (КФ 2.5.1.6) при получении [2H]серина [154].
Ферментативный метод давно используется для препаративного лабораторного и промышленного получения оптически активных аминокислот, благодаря высокой субстратной специфичности ферментов и возможности селективного введения стабильных изотопов по определённым положениям молекул аминокислот. Основными аспектами использования ферментативных систем являются каталитические реакции ассиметрического образования связи на прохиральных субстратах и ферментативное разделение рацематов аминокислот.
Что касается хроматографического разделения рацематов на прохиральных сорбентах, то оно все же недостаточно эффективно для разделения и обеспечивает в лучшем случае больше половины меченого продукта в виде одного из оптических антиподов. Ферментативная стадия часто завершает химический синтез меченых аналогов аминокислот, причем использование для этих целей интактных клеток или их экстрактов так же эффективно, как использование очищенных ферментов. Однако, субстратная специфичность ферментов, их ограниченная доступность, сложность их выделения и очистки ограничивают их применение для этих целей. Несмотря на то, что ферментативные синтезы преодолевают все вышеперечисленные проблемы, низкие выхода очищенных ферментов лимитируют использование химико-ферментативных реакций. Так, разработанный ферментативный процесс для получения [15N]аланина включает комплексное использование нескольких специфичных ферментов и меченых субстратов и имеет выход по целевому продукту не более 1 г [155]. С другой стороны, методы генной инженерии открывают возможности для получения большинства ферментных препаратов в препаративных количествах.
Методы получения [15N]аминокислот связаны с использованием [15N]аммонийных или [15N]нитратных солей в качестве источников 15N-метки [156], в то время как ферментативный метод более эффективен для получения прежде всего [15N]аспарагиновой и [15N]глутаминовой кислот за счёт аминирования -кетопроизводных аминокислот и в тех случаях, когда необходимы высокие уровни включения изотопа 15N в молекулы [157].
Осуществление различных методов включения 15N-метки в молекулы аминокислот связано с использованием методов газовой подпитки 15NН3 [158], иммобилизацией клеток с последующей активацией носителя 15NН4Cl [159], или с оптимизацией концентраций [15N]предшественников аминокислот в ростовых средах [160]. C использованием вышеперечисленных подходов были получены [15N]аспарагиновая кислота и [15N]аланин с уровнями изотопного включения 15N, превышающими 95% [161]. При этом иммобилизованные клетки E. coli были использованы как источник аспартазы, которая катализирует превращение [15N]фумаровой кислоты и [15N]фумарата в [15N]аспарагиновую кислоту, в то время как для получения [15N]аланина из [15N]аспарагиновой кислоты в качестве источника аспартат-4-декарбоксилазы использовали бактерию Pseudomonas decahee. [15N]глутаминовую кислоту получали за счёт процесса ферментации бактерий Brevibacterium lactofermentum с предшественниками аминокислот в присутствии 15NН4ОН [162].
Для включения изотопа 13С в молекулы аминокислот могут применяться аналогичные ферментативные подходы с применением [13С]глюкозы, однако при проведении ферментативной реакции требуются значительные количества высокообогащённой [13C]глюкозы и поэтому даный метод является слишком дорогим для получения [13C]аминокислот. Кроме того, большая часть глюкозы (до 70%) идёт на обеспечение процесса дыхания клетки, поэтому эффективность мечения молекул БАС изотопом углерода за счёт ферментативного окисления [13С]глюкозы невысокая. Так, уровни изотопного обогащения [13C]глутамата, полученного ферментативно с участием [13C]глюкозы, были менее 50% [163].
Перспективны также подходы с использованием комбинации химико-ферментативных и биотехнологических способов получения изотопномеченых аминокислот. В работах [164, 165] сообщается о получении более десятка аналогов триптофана, специфически меченных изотопами 2Н, 13C, 15N по индольному кольцу молекулы. Моно-изотопномеченые производные индолов и их 4, 5 и 7-гидроксипроизводные были ферментативно превращены в меченые аналоги триптофана при помощи генетически сконструированного штамма бактерий E. coli, содержащего рекомбинантную плазмиду с триптофановым (Тrp) опероном, который кодировал ряд ферментов, ответственных за биосинтез этой аминокислоты.
В заключение следует отметить, что несмотря на многочисленность описанных в современной литературе подходов по получению БАС, меченных стабильными изотопами, в настоящее время практически не существует способов, которые позволяют получать аминокислоты и белки, меченные 2Н, 13С, 15N и 18O за счет того или иного универсального подхода, хотя химико-ферментативные методы позволяют использовать одну и ту же химико-биохимическую реакцию для получения меченых аминокислот за счет применения различных меченых низкомолекулярных реагентов (субстратов). Получение аминокислот и белков, высокообогащённых стабильными изотопами удобнее всего проводить с использованием биотехнологических подходов, в то время как селективности включения стабильных изотопов в молекулы БАС можно достичь за счёт применения комбинации синтетических и ферментативных реакций. Выбор метода получения БАС, несущих ту или иную изотопную метку, определяется прежде всего целью исследования.
ЛИТЕРАТУРА.
1. Smith K., Barua J. M., Watt P. W. // Am. J. Physiol. - 1992. - V. 262. - N 3. - P. 1372-1376.
2. McIntosh L. P., Dahlquist F. W. // Quarterly Reviews of Biophysics. - 1990. - V. 23. - N. 1. - P. 1-38.
3. Harbison G. S., Smith S. O., Pardoen J. A., Mulder P. P. J., Lugtenburg J., Herzfeld R., Mathies R., Griffin R. G. // Biochemistry. - 1984. - V. 23. - P. 2662-2667.
4. Ames J. B., Ros M., Raap J., Lugtenburg J., Mathies R. A. // Biochemistry. - 1992. - V. 31. - P. 5328-5335.
5. Fischer M. R., de Groot H. J. M., Raap J., Winkel C., Hoff A. J., Lugtenburg J. // Biochemistry. - 1992. - V. 31. - P. 11038-11043.
6. Lewis A., Marcus M. A., Ehrenberg B., Crespi H. L. // Proc. Nat. Acad Sci. USA. - 1978. - V. 75. - P. 4642-4646.
7. Fesik S. W., Eaton H. L., Olejniczak E. T., Zuiderweg E. R. P., McIntosh L. P., Dahlquist F. W. // J. Am. Chem. Soc. - 1990. - V. 112. - P. 886-888.
8. Motil K. J., Montandon C. M., Thotathuchery M., Garza C. // Am. J. Clin. Nutr. - 1990. - V. 51. - P. 378-384.
9. Young V. R., Yu Y. M., Krempf M. Protein and amino acid turnover using the stable isotopes 15N, 13C, and 2H as probes. New techniques in nutritional research // Eds. R. G. Whitehead, A. J. - Prentice. - Academic Press. - New York, 1990. - V. 9. - P. 17-72.
10. Digenis G. A., Goto R., Chaney J. E., Tamemasa O. // J. Pharm. Sci. - 1982. - V. 71. - P. 816-820.
11. Nelson J. E., Ruo T. I. // Clinica Chemica Acta. - 1988. - V. 175. - P. 59-65.
12. Beaufrere B., Fournier V., Salle B., Putet G. // Am. J. Physiol. - 1992. - V. 263. - N 2. - P. 214-220.
13. Darmaun D., Robert J. J., Bier D. M., Matthews D. E., Young V. R. // Annales-d’Endocrinologie. - 1985. - V. 46. - N 4. - P. 355-356.
14. Hruby V. J. // Synth. and Appl. Isot. Label. Compounds. - 1985. - V. 4. - P. 287-292.
15. Viswanatha V., Hruby V. J. // J. Org. Chem. - 1980. - V. 45. - P. 2010-2015.
16. Fesik, S. W., Zuiderweg, E. R. P.// Quarterly Reviewes of Biophysics. - 1990. - V. 23. - N 2. - P. 97-131.
17. Ellman J. A., Volkman B. F., Mendel D. // J. Am. Chem. Soc. - 1992. - V. 114. - P. 7959-7961.
18. Griesinger C., Sorensen O. W., Ernst R. R. // J. Am. Chem. Soc. - 1987. - V. 109. - P. 7227-7228.
19. Zuiderweg E. R. P., McIntosh L. P., Dahlquist F. W., Fesik S. W. // J. Magn. Reson. - 1990. - V. 86. - P. 210-216.
20. Rothschild, K. J., Braiman, M. S., He, Yi-Wu., Marti, T., Khorana, H. G. // J. of Biol. Chem. - 1990. - V. 28. - P. 16985-16991.
21. Haris P. I., Robillard G. T., Vandijk A. A., Chapman D. // Biochemistry. - 1992. - V. 31. - N 27. - P. 6279-6284.
22. Argade, P. V., Rothschild, K. J., Kawamoto, A. H., Herzfeld, J., Herlihy, W. C. // Proc. Natl. Acad. Sci. USA. - 1981. - V. 78. - N 3. - P. 1643-1646.
23. Eisenstein L., Lin S., Dollinger G., Odashima K., Termini J., Konno K., Ding W., Nakanischi K. // J. Am. Chem. Soc. - 1987. - V. 109. - P. 6860-6862.
24. Irving C. S., Nissim I., Lapidot A. // Biomed. Mass. Spectrom. - 1978. - V. 5. - P. 117-122.
25. Raap J., Winkel C., de Wit A. H. M., van Houten A. H. H., Hoff A. J., Lugtenburg J. // Anal. Biochem. - 1990. - V. 191. - P. 9-18.
26. Мосин О. В., Егорова Т. А., Складнев Д. А., Швец B. И. // Биоорганическая химия. - 1996. - Т. 22. - N 10-11. - С. 861-874.
27. Мосин О. В. Разработка методов биотехнологического получения белков, аминокислот и нуклеозидов, меченных 2Н (D) и 13С, с высокими степенями изотопного обогащения. Автореф. дис. канд. хим. наук. М.: МИТХТ им. М. В. Ломоносова. - 1996. - С. 3-23.
28. Мосин О. В., Егорова Т. А., Чеботаев Д. А., Складнев Д. А., Юркевич А. М., Швец В. И. // Биотехнология. - 1996. - N 4. - С. 27-34.
29. LeMaster D. M. // Quarterly Reviews of Biophysics. - 1990. - V. 23. - N. 1. - P. 133-174.
30. Пшеничникова А. Б., Карнаухова Е. Н., Звонкова Е. Н., Швец В. И. // Биоорганическая химия. - 1995. - Т. 21. - N 3. - С. 163-178.
31. Raap J., van der Wielen C. M., Lugtenburg J. // Recl. Trav. Chim. Pays-Bas. - 1990. - V. 109. - P. 277-286.
32. Winkel C., Aarts M. W. M., van der Heide F. R., Buitenhuis E. G., Lugtenburg J. // Recl. Trav. Chim. Pays-Bas. - 1989. - V. 108. - P. 139-146.
33. Raap J., Wolthuis W. N. E., Hehenkamp J. J. J., Lugtenburg J. // Amino Acids. - 1995. - V. 8. - P. 171-186.
34. van den Berg E. M. M., Baldew A. U., de Goede A. T. J., Raap J., Lugtenburg J. // Recl. Trav. Chim. Pays-Bas. - 1988. - V. 107. - P. 73-81.
35. van den Berg E. M. M., Richardson E. E., Lugtenburg J. // Synth. Comm. - 1987. - V. 17. - N 10. - P. 1189-1198.
36. Ragnarsson U. // Journal of Peptide Science. - 1995. - V. 3. - P. 149-156.
37. Cappon J. J., van der Walle G. A. M., Verdegem P. J. E., Raap J., Lugtenburg J. // Recl. Trav. Chim. Pays-Bas. - 1992. - V. 111. - P. 517-523.
38. Loftfild R. B., Eigner E. A. // Biochim. Biophys. Acta. - 1966. - V. 130. - P. 449-457.
39. Lugwig S. N., Unkefer C. J. // J. Labelled Compd. Radiopharm. - 1992. - V. 31. P. 95-102.
40. Berger A., Smolarsky M., Kurn N., Bosshard H. R. // J. Org. Chem. - 1973. - V. 38. - P. 457-460.
41. Daub G. H. Syntheses with stable isotopes. Stable Isotopes. Proc. of the 3d Intern. Conference. // Ed. E. R. Klein. - Academic Press. - New York, 1979. - P. 3-10.
42. Jones W. C., Rothgeb T. M., Gurd F. R. N. // J. Biol. Chem. - 1976. - V. 251.- P. 7452-7460.
43. Sternlicht H., Kenyon G. L., Packer E. L., Sinclair J. // J. Am. Chem. Soc. - 1971. - V. 93. - P. 199-208.
44. Giza Y. H., Ressler C. // J. Labelled Compd. - 1969. - V. 5. - P. 142-151.
45. Havranek M., Kopecka-Schadtova H., Veres K. // J. Labelled Compd. - 1970. - V. 6. - P. 345-354.
46. Wilcox M. // Anal. Biochem. - 1974. - V. 59. - P. 436-440.
47. Matthews H. R., Matthews K. S., Opella S. J. // Biochim. et Biophys. Acta. - 1977. - V. 497. - P. 1-13.
48. Bak B., Dambmann C., Nicolaisen F., Pederson E. J., Bhacca E. J. // J. Mol. Spectrosc. - 1968. - V. 26. - P. 78-97.
49. Griffiths, D. V., Feeney, J., Roberts, G. C. K., Burgen, A. S. V. // Biochim. et Biophys. Acta. - 1976. - V. 446. - P. 479-485.
50. Kinsey, R. A., Kintanar, A., Oldfield, E. // J. Biol. Chem. - 1981. - V. 256. - N 17. - P. 9028-9036.
51. Золотарёв Ю. А., Зайцев Д. А., Татур В. Д., Мясоедов Н. Ф. Способ получения равномерно меченных дейтерием оптически активных -аминокислот: А. с. N 1685903 СССР. - // ВНИИГПЭ. - 1991. - N 39. - С. 92.
52. Золотарёв Ю. А., Козин В. С., Зайцев Д. А., Дорохова Е. Н., Мясоедов Н. Ф. // Докл. АН СССР. - 1989. - Т. 308. - N 5. - С. 1146-1150.
53. Samuel D. Methology of oxygen isotopes. Oxygenases. // Ed. M. Hayaishi. - Academic Press. - New York, 1972. - P. 31-86.
54. Bunton C. A., James D. H., Senior J. B. // J. Chem. Soc. - 1960. - P. 3364-3367.
55. Busujima U. K., Shimiba S., Narita K., Okada S. // Chem. Pharm. Bull. - 1988. - V. 36. - P. 1828-1832.
56. Lapidot A., Kahana Z. E. // Trends Biotechnol. - 1986. - V. 4. - P. 2-4.
57. McIntosh L. P., Griffey R. H., Muchmore D. C., Nielson C. P., Redfield A. G., Dahlquist F. W. // Proc. Natn. Acad. Sci. USA. - 1987. - V. 84. - P. 1244-1248.
58. Westler W. M., Kainosho M., Nagao H., Tomonaga N., Markley J. L. // J. Am. Chem. Soc. - 1988. - V. 110. - P. 4093-4095.
59. Westler W. M., Stockman B. J., Markley J. L., Hosoya Y., Miyake Y., Kainosho M. // J. Am. Chem. Soc. - 1988. - V. 110. - P. 6256-6258.
60. Katz J., Crespi H. L. // Pure Appl. Chem. - 1972. - V. 32. - P. 221-250.
61. Murphy R. C., Anderson F. S., Clay K. L. In vitro and in vivo studies of amino acids labeled with 18O at the carboxyl moiety. Stable Isotopes. Proc. of the 3d Intern. Conference. // Ed. E. R. Klein. - Academic Press. - New York, 1979. - P. 139-146.
62. Cox J., Kyli D., Radmer R. // Trends Biotechnol. - 1988. - V. 6. - P. 279-282.
63. Егорова Т. А., Мосин О. В., Ерёмин С. В., Карнаухова Е. Н., Звонкова Е. Н., Швец В. И. // Биотехнология. - 1993. - N 8. - С. 21-25.
64. Katz J. J., Crespi H. L. // Science. - 1966. - V. 151. - P. 1187-1194.
65. Tran-Dinh S., Fermandjian S., Sala E., Mermet-Bouvier R., Cohen M., Fromageot P. // J. Am. Chem. Soc. - 1974. - V. 96. - P. 1484-1493.
66. Oh B. H., Westler W. M., Darba P., Markley J. L. // Science. - 1988. - V. 240. - P. 908-911.
67. Stockman B. J., Reily M. D., Westler W. M., Ulrich E. L., Markley J. L. // Biochemistry. - 1989. - V. 28. - P. 230-236.
68. Yu L. P., Smith G. M. // Biochemistry. - 1988. - V. 27. - P. 1949-1956.
69. Stockman B. J., Westler W. M., Mooberry E. S., Markley J. L. // Biochemistry. - 1988. - V. 27. - P. 136-142.
70. McInnes A. G., Walter J. A., Wright J. L. C., Vining L. C. 13C NMR biosynthetic studies. Topics in carbon- С13NMR spectroscopy. //Ed. G. D. Levy. - John Wiley & Sons, Inc. - New York, 1976. - V. 2. - P. 125-178.
71. Weber P. L., Muller L. // J. Magn. Res. - 1989. - V. 81. - P. 430-434.
72. Crespi H. L. Biosynthesis and uses of per-deuterated proteins. Synt. and Appl. of Isot. Label. Compd. // Ed. R. R. Muccino. - Elsevier. - Amsterdam, 1986 - P. 111-112.
73. Мосин О. В., Карнаухова Е. Н., Пшеничникова А. Б., Складнев Д. А., Акимова О. Л. // Биотехнология. - 1993. - N 9. - С. 16-20.
74. Daboll H. F., Crespi H. L., Katz J. J. // Biotechnology and Bioengineering. - 1962. - V. 4. - P. 281-297.
75. Crespy H. L. Stable Isotopes in the Life Sciences. // 2nd edn. - International atomic energy agency. - Vienna, 1977. - P. 111-121.
76. Matveev A. V., Mosin O. V., Skladnev D. A., Yurkevich A. M. // Methylotrophic adaptation to highly deuterated substrates. Proc. 8th Int. Symp. Microb. Growth on C1-Compounds. - 27 August 1995. - San Diego. - U.S.A. - P. 80.
77. Мосин О. В., Казаринова Л. А., Преображенская Е. С., Складнев Д. А., Юркевич А. М., Швец В. И. // Биотехнология. - 1996. - N 4. - С. 19-26.
78. Казаринова Л. А., Королькова Н. В., Миронов А. С., Мосин О. В., Складнев Д. А., Юркевич А. М. Способ получения высокодейтерированных нуклеозидов и нуклеотидов. - Заявка РФ N 95118778 от 14.11.1995.
79. Brazier I. L., Ridon B., Falconnet I. B., Charrah V., Benchekrour Y. // Therapie. - 1987. - V. 42. - N 5. - P. 445-450.
80. Sharon N., Grisar V., Newmann H. // Arch. Biochem. Biophys. - 1962. - P. 219-221.
81. Randall M., Kathleen S. M., Stanley J. O. // Biochim. Biophys Acta. - 1977. - V. 497. - P. 1-5.
82. Beeney J., Birdsall B., Oster G., Carr M. D., Karine M. A. // FEBS Lett. - 1990. - V. 272. - ¹ 1. - P. 197-199.
83. Umbarger H. E. // Ann. Rev. Biochem. - 1978. - V. 47. - P. 533-606.
84. Bachmann, B. J. // Microbiol. Rev. - 1983. - V. 47. - P. 180-230.
85. Griffey R. H., Redfield A. G., Loomis R. E., Dahlquist F. W. // Biochemistry. - 1985. - V. 24. - P. 817-822.
86. Bachovchin W. M. // Proc. Natl. Acad. Aci. - 1985. - V. 82. - P. 7948-7951.
87. Bachovchin W. M. // Biochemistry. - 1986. - V. 25. - P. 7751-7759.
88. Patel G. B., Roth L. A. // Can. J. Microbiol. - 1978. - V. 24. - P. 1007-1010.
89. Wood H. G., Ragsdale S. W., Pezacka E. // Trends Biochem. Sci. - 1986. - V. 11. - P. 14-18.
90. Daniels L., Sparling R., Sprott G. D. // Biochim. Biophys. Acta. - 1984. - V. 768. - P. 113-163.
91. Fukuzaki S., Nishio N., Nagai S. // Appl. and Environ. Microbiol. - 1990. - V. 56. - P. 3158-3163.
92. Patel G. B., Sprott G. D., Fein J. E. // Int. J. Syst. Bacteriol. - 1990. - V. 40. - P. 79-82.
93. Ekiel I., Smith I. C. P., Sprott G. D. // J. Bacteriol. - 1983. - V. 156. - P. 316-326.
94. Ekiel I., Jarrell K. F., Sprott G. D. // Eur. J. Biochem. - 1985. - V. 149. - P. 437-444.
95. Fuchs G., Stupperich E. // Arch. Microbiol. - 1980. - V. 127. - P. 267-272.
96. Jansen K., Stupperich E., Fuchs G. // Arch. Microbiol. - 1980. - V. 132. - P. 355-364.
97. Bender D. A. Amino Acid Metabolism. // 2nd edn. - New York. - John Wiley & Sons, 1985. - P. 34-52.
98. Charon N. W., Johnson R. C., Peterson D. // J. Bacteriol. - 1974. - V. 117. - P. 203-211.
99. Kisumi M., Komatsubara S., Chibata I. // J. Biochem. - 1977. - V. 82. - P. 95-103.
100. Vollbrecht D. // Biochim. Biophys. Acta. - 1974. - V. 362. - P. 382-389.
101. Conn E. E. // The Shikimic Acid Pathway. Recent Advances in Phytochemistry. - Plenum Press. - New York, 1986. - V. 20. - P. 34-40.
102. McFadden B. A. Assimilation of One-Carbon Compounds. The Bacteria, a Treatise on Structure and Function. // Ed. L. N. Ornston. - Academic Press. - New York, 1978. - V. 4. - P. 219-290.
103. Никонова Е. С., Доронина Н. В., Троценко Ю. А. // Приклад. биохим. и микробиол. - 1986. - Т. 22. - С. 557-561.
104. Hanson R. S., Dillingham R., Olson P., Lee G. H., et all. Production of L-lysine and some other amino acids by mutants of B. methanolicus. Microbial Growth on C1 Compounds. // Ed. M. E. Lindstrom. - Kluwer Academic Publishers, 1996. - P. 227-236.
105. Anthony C. Bacterial oxidation of methane and methanol. Advances in Microbial Physiology. // Eds. Rose A. H., Tempest D. W. - Academic Press. - New York, 1986. - V. 27. - P. 113-203.
106. Higgins I. J., Quayle J. R. // Biochemical Journal. - 1970. - V. 118. - P. 201-210.
107. Goldberg I. // Eur. J. Biochem. - 1976. - V. 63. - P. 233-239.
108. Dalton H. // Advances in Applied Microbiology. - 1980. - V. 26. - P. 71-80.
109. Colby J., Dalton H., Whittenbury R. // Ann. Rev. Microbiol. - 1979. - V. 33. - P. 481-517.
110. Kemp M. B., Quayle J. R. // Biochem J. - 1967. - V. 102. - P. 94-102.
111. Mosin O. V., Karnaukhova E. N., Skladnev D. A. // Application of methylotrophic bacteria for preparation of stable isotope labelled amino acids. Proc. 7th Int. Symp. Genetics of Industrial Microorganisms. - Quebec. Canada. - 26 June 1994. - P. 163.
112. Mosin O. V., Karnaukhova E. N., Skladnev D. A. // Preparation of 2H- and 13C-amino acids via bioconvertion of C1-substrates. Proc 8th Int. Symp. Microb. Growth on C1-Compounds. - San Diego. U.S.A. - 27 August 1995. - P. 80.
113. Мосин О. В., Карнаухова Е. Н., Складнев Д. А., Акимова О. Л., Цыганков Д. Ю. Штамм Brevibacterium methylicum - продуцент униформно меченой дейтерием аминокислоты - L-фенилаланина. Заявка РФ N 93055824. // ВНИИГПЭ. - 1993. - N 94. - С. 3-4.
114. Karnaukhova, E. N., Reshetova, O. S., Semenov, S. Y., Skladnev, D. A., and Tsygankov, Y. D. // Amino Acids. - 1994. - V. 6. - P. 165-176.
115. Karnaukhova E. N., Mosin O. V., Reshetova O. S. // Amino Acids. - 1993. - V. 5. - P. 125.
116. Складнев Д. А., Мосин О. В., Егорова Т. А., Ерёмин С. В., Швец В. И. // Биотехнология. - 1996. - N 5. - С. 14-20.
117. Мосин О. В., Складнев Д. А., Егорова Т. А., Юркевич А. М., Швец В. И. // Биотехнология. - 1996. - N 3. - С. 3-12.
118. Muchmore D. C., McIntosh L. P., Russel C. B., Anderson E. E., Dahlquist F. W. // Meth. Enzymol. - 1990. - V. 77. - P. 346-354.
119. DeVault J. D., Hughes K. J., Johnson O. A., Narang S. K. // Biotechnology. - 1996. - V. 14. - P. 46-49.
120. Torchia D. A., Sparks S. W., Bax A. // Biochemistry. - 1989. - V. 28. - P. 5509-5524.
121. Marion D., Kay L. E., Sparks S. W., Torchia D. A., Bax A. // J. Am. Chem. Soc. - 1989. - V. 3. - P. 1515-1517.
122. Senn H., Euguster A., Otting G., Suter F., Wuthrich K. // Eur. Biophys. J. - 1987. - V. 14. - P. 301-313.
123. LeMaster D. M., Richards F. M. // Biochemistry. - 1985. - V. 24. - P. 7263-7268.
124. Lapidot, A., Irving, C. S. // J. Am. Chem. Soc. - 1977. - V. 99. - P. 5488-5493.
125. Bachovchin W. M., Roberts J. D. // J. Am. Chem. Soc. - 1978. - V. 100. - P. 8041-8046.
126. Kainosho M., Tsuji T. // Biochemistry. - 1982. - V. 21. - P. 6273-6279.
127. Leighton P., Lu P // Biochemistry. - 1987. - V. 26. - P. 7262-7271.
128. Campbell B. S., Papastavros M. Z., McCormick F., Redfield A. G. // Proc. Natl. Acad. Sci. USA. - 1989. - V. 86. - P. 817-820.
129. Skladnev D. A., Tsygankov Y. D. // Conversion of stable isotope labeled methanol to components of bacterial biomass. Proc. 6 th Eur. Conf. on Biomass for Energy. - Athens. - Greece. - 22-26 April. - 1991. - P. 234.
130. Fesik S. W., Eaton H. L., Olejniczak E. T., Zuiderweg E. R. P., McIntosh L. P., Dahlquist F. W. // J. Am. Chem. Soc. - 1990. - V. 112. - P. 886-888.
131. Gelfand D. H., Steinberg R. A. // J. Bacteriol. - 1977. - V. 130. - P. 429-440.
132. LeMaster D. M., Cronan J. E. // J. of Biological Chemistry. - 1982. - V. 257. - N 3. - P. 1224-1230.
133. Ниедервайзер А., Патаки Д. Новые методы анализа аминокислот, пептидов и белков. - М.:Мир, 1974. - С. 20-73.
134. Овчинников Ю. А. Биоорганическая химия. - М.: Просвещение, 1987. - С. 41-48
135. Spande T. F., Witkop B., Degani Y., Patchornik A. // Adv. Protein Chem. - 1970. - V. 24. - P. 97-260.
136. Hill R. L. // Adv. Protein Chem. - 1965. - V. 20. - P. 37-107.
137. Moore S. Chemistry and Biology of Peptides // Ed. J. Meienhofer. - Ann Arbor Sci. Publ. - Ann Arbor. - Michigan, 1972. - P. 629-653.
138. Muramoto K., Kamiya H. // Anal. Biochem. - 1990. - V. 189. - P. 223-230.
139. Matsubara H., Sasaki R. M. // Biochim. Biophys. Res. Com. - 1969. - V. 35. - P. 175-177.
140. Ng L. T., Pascaud A., Pascaud M. // Anal. Biochem. - 1987. - V. 167. - P. 47-52.
141. Пшеничникова А. Б., Карнаухова Е. Н., Мицнер Б. И. // Ж. общ. химии. - 1993. - Т. 63. - Вып. 5. - С. 1034-1040.
142. Bak B., Led J. J., Pederson E. J. // Acta Chem. Scand. - 969. - V. 23. - P. 3051-3054.
143. LeMaster D. L., Richards F. M. // Anal. Biochem. - 1982. - V. 122. - P. 238-247.
144. Cohen J. S., Putter I. // Biochim. et Biophys. Acta. - 1970. - V. 222. - P. 515-520.
145. Patel G. B., Sprott G. D., Ekiel I. // Appl. Environ. Microbiol. - 1993. - P. 1099-1103.
146. Егорова Т. А., Еремин С.В ,Мицнер Б.И, Звонкова Е.. Н., Швец В.И. Биотехнология. - 1993. - N 5. - С. 32-36.
147. Egorova T. A., Eremin S. V., Mitsner B. I., Zvonkova E. N., Shvets V. I. // J. of Chromatography B. - 1995. - V. 665. - P. 53-62.
148. Prescan E., Ivanof A., Mocanu A., Palibroda N., Bologa M., Gorun V., Barzu O. // Microbiol. Technol. - 1987. - V. 9. - P. 663-665.
149. Hadener A., Tamm C. // J. Labelled Compounds and Radiopharm. - 1987. - V. 24. - P. 1291-1306.
150. Walker T. E., Matheny C. // J. Org. Chem. - 1986. - V. 51. - P. 1175-1179.
151. Fuganti C., Ghiringhelli D., Giangrasso D., Grasselli P. // J. Chem. Soc. Comm. - 1974. - P. 726-730.
152. Сappon J. J., Baart J., van der Walle G. A. M., Raap J., Lugtenburg J. // Recl. Trav. Chim. Pays-Bas. - 1991. - V. 110. - P. 158-166.
153. Field S. J., Young D. W. // J. Chem. Soc. Chem. Communs. - 1979. - P. 1163-1165.
154. Jordan P. H., Akhtar H. // J. Biochem. - 1970. - V. 116. - P. 277-286.
155. Greenway W., Whatley F. R., Ward S. // Febs Lett. - 1977. - V. 81. - P. 286-288.
156. Yamada S., Nabe K., Izuo N., Nakamichi K., Chibata I. // Appl. Environ. Microbiol. - 1981. - V. 42. - P. 773-778.
157. Hirose Y., Shibai H. // Biotechnol. Bioeng. - 1980. - V. 22. - Suppl. 1. - P. 111-125.
158. Nabe K., Ujimaru T., Izuo N., Yamada S., Chibata I. // Appl. Environ. Microbiol. - 1980. - V. 40. - P. 19-24.
159. Chibata I., Tosa T., Sato T. Microbial technology. // Ed. H. J. Peppler. - 2nd ed. Academic Press, 1979. - V. 11. - P. 433-461.
160. Kahana Z. E., Lapidot A. Proc. 3d European Congress of Biotechnology. - Verlag-Chemie. - Weinheim, 1984. - V. 1. - P. 439-443.
161. Kahana Z. E., Lapidot A. // Anal. Biochem. - 1982. - V. 126. - P. 389-393.
162. Kahana Z. E., Lapidot A. // Anal. Biochem. - 1983. - V. 132. - P. 160-164.
163. Kahana Z. E., Lapidot A. Adaptation of biotechnological processes for preparation of compounds labeled with stable isotopes. Synthesis and Applications of Isot. Label. Compounds. // Ed. R. R. Muccino. - Elsevier, 1986. - P. 511-512.
164. van der Berg E. M. M.. Synthesis of isotopomers of L-tryptophan using genetically modified Escherichia coli bacteria. - Ph. D. Dissertation. - University of Leiden. - The Netherlands, 1989. - P. 1-123.
165. van der Berg E. M. M., van Liemt J. H., Willem B. S. // Recl. Trav. Chim. Pays-Bas. - 1989. - V. 108. - N 9. - P. 304-313.
THE METHODS FOR PRODUCTION OF AMINO ACIDS AND PROTEINS LABELLED WITH STABLE ISOTOPS 2H, 13C, 15N, 18O
O. V. Mosin
M. V. Lomonosov State Academy of Fine Chemical Technology, Vernadskogo Prospect 86, Moscow, 117571
This given review is devoted to development of modern biotechnological and chemico-phermentation methods on production of amino acids and proteins labelled with stable isotopes 2Н, 13С, 15N, 18О. Potential opportunities of these methods for the directed synthesis of isotopically labelled amino acids and proteins are considered. There are submitted data on preparation and further using of synthesized isotopically labelled compounds for various biochemical studies with application of methods of spectroscopy of a nuclear magnetic resonance, infra-red and mass-spectrometry.