
Текст взят с психологического сайта
| Вид материала | Документы |
СодержаниеИндукция как модель хранения следов памяти Консолидация следов Затухание активности в нервной системе и тормозные взаимодействия |
- Текст взят с психологического сайта, 6189.05kb.
- Текст взят с психологического сайта, 4254.71kb.
- Текст взят с психологического сайта, 1854.21kb.
- Текст взят с психологического сайта, 11863.68kb.
- Текст взят с психологического сайта, 8514.9kb.
- Текст взят с психологического сайта, 3673.56kb.
- Текст взят с психологического сайта, 8427.66kb.
- Текст взят с психологического сайта, 8182.42kb.
- Текст взят с психологического сайта, 5461.28kb.
- Текст взят с психологического сайта, 5587.31kb.

так и после тренировки крыс. С помощью этой техники было обнаружено, например, что вещество, которое препятствует синтезу протеинов, может нарушать процесс припоминания, если лнъекция сделана спустя 5 или 6 часов после того, как крыса приобрела опыт в выполнении задания. Это означает, что эти вещества вызвали блокаду синтеза протеинов. Однако позднее было показано, что, если сохранение следов проверяется через несколько недель после инъекции, припоминание задания ухуд-
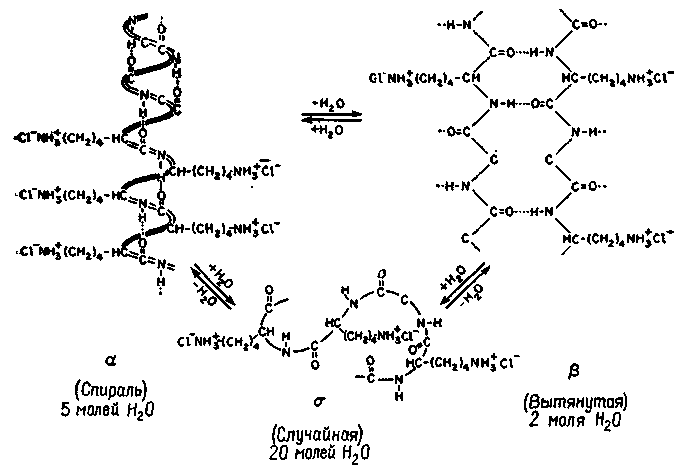
Рис. 11-11. Схема возможных конфигураций молекулярных структур, которые может принимать молекула нуклеиновой кислоты (полимер poly-L-lysine hydrochlorid) (Blout, 1967).
«пается лишь незначительно. Инъекция этих веществ, по-видимому, нарушает не конструкцию молекулы памяти, а, скорее, препятствует процессу обращения к ней (Agranoff, Davis and Brink, 1965). Однако роль протеинов и других макромолекул (особенно ли-пидов мозга) в механизме памяти, возможно, не столь ограниченна, как это предполагается гипотезой, проверяемой экспериментами с введением веществ", тормозящих обмен. В силу своей сложности макромолекулы имеют уникальные конфигурации, -структуры, которые могут меняться на короткое время и вызывать другое состояние (рис. II-11). С помощью структурных изменений можно хорошо объяснить временную память, связанную с такими образами, которые характерны для лиц с «фотографической» (или «эйдетической») памятью. Один такой эйдетиче-
54
ски одаренный студент недавно был обнаружен в Гарварде. В результате экспериментальной проверки было показано, что он способен сохранять в памяти каждую деталь своих зрительных восприятий в течение 8 дней. Вторая часть книги будет в значительной мере посвящена рассмотрению этой временной спирализации молекул. Изучение структурных изменений мозговой ткани сейчас едва ли возможно, однако техника развивается такими темпами, что эта область исследования многое обещает уже в ближайшем будущем (Sjostrand, 1969).
Некоторые исследователи продолжают придерживаться гипотезы, что РНК является непосредственным хранилищем памяти, несмотря на серьезные доводы против этой гипотезы. Они основывают свою точку зрения яа весьма спорных экспериментах с «переносом», которые были проведены на планариях, крысах и обезьянах. В этих экспериментах экстракт РНК от контрольных животных и животных, обучавшихся выполнению задания, вводили необученным животным. Те из них, которым вводили РНК от животных, имевших опыт обучения, иногда, но не всегда решали задачи быстрее по сравнению с теми, которым вводили РНК от контрольных животных. Согласно некоторым данным, эффект различного действия РНК «обученного» и «контрольного» животного исчезал в результате обработки экстракта РНК веществом, избирательно разрушающим РНК (см. обзор McGonnell, 1970).
Сторонники химической гипотезы сохранения следов, оказавшись перед необходимостью выбора между двумя возможными решениями, в конце концов остановились на нейронных медиаторах. Уменьшение скорости и амплитуды нервных импульсов, когда они достигают окончаний аксонов, возникает из-за заметного уменьшения диаметра окончаний волокон. Следовательно, пресинаптических потенциалов самих по себе еще далеко не достаточно, чтобы вызвать постсинаптический потенциал. Однако пресинаптического электрического заряда достаточно, чтобы дать-толчок высвобождению медиатора, который хранится на оконча-
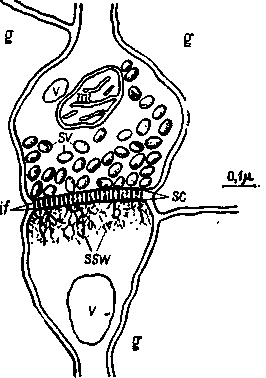
Рис. И-12. Типичная клетка коры. Обычная форма синапса-в мозгу млекопитающих. Ак-сонная (пресинаптическая) сторона вверху; дендритная (постсинаптическая) сторона — внизу, g — рлия; if — внутрисинаптические волоконца; mi — митохондрия, sc — синаптическая щель; ssw — субсинаптическая паутина; sv — синэптические пузырькн; v — везикулярное тело (Calvin. 1967, см. Roberts, 1S62).
55
нии аксона в маленьких пузырьках (рис. П-12). Память могла бы зависеть от легкости, с которой высвобождается медиатор, или от его количества. Таким образом, можно провести такие эксперименты — и они действительно проводятся, — когда медиатор нейтрализуется в результате действия фармакологических веществ или его разрушение блокируется во время тренировки животного (Deutsch, Hamburg and Dahl, 1966). В результате таких действий у крыс нарушается процесс научения, но, как и в исследованиях с применением веществ, тормозящих обмен, возникает вопрос, не вызывает ли введение фармакологических веществ побочных эффектов, меняющих деятельность мозга и, следовательно, влияющих на процесс запоминания, — таких побочных эффектов, как появление локальных электрических разрядов в частях мозга, не связанных непосредственно с сохранением следов памяти, как таковой, но препятствующих обращению к ней во время проверки результатов научения.
ИНДУКЦИЯ КАК МОДЕЛЬ ХРАНЕНИЯ СЛЕДОВ ПАМЯТИ
Моя реакция на изобилие экспериментальных данных, полученных при исследовании энграммы в 1960-е гг., двойственна. Во-первых, можно прийти к выводу, что существует не один механизм памяти, не единая молекула памяти. Пластичность нейрона многообразна, и память не является единым процессом. Воображение, узнавание, припоминание предполагают использование некоторых или всех основных форм пластичности нервной ткани и различных способов сохранения и воспроизведения следа. Поэтому первый вывод, который следует сделать, состоит в том, что память, как она обычно представляется, не единый механизм, лежащий в основе процессов, которые позволяют организму устанавливать связи между отдельными звеньями опыта.
Во-вторых, простая модель следов, предполагающая постоянную модификацию мозговой ткани на нейронном уровне, может быть сформулирована уже сейчас. Такая модель должна обеспечить не только возможность сохранения следов, но и доступность считывания зафиксированного изменения. Наконец, такая модель должна быть основана на уже имеющихся данных и отвечать здравому биологическому смыслу. |J3 настоящее время наиболее вероятным источником такой модели может служить аналогия с процессом, происходящим во время эмбрионального развития организма (рис. П-13). Такие структуры, как глаз, формируются только тогда, когда ткань, из которой они развиваются, соответствующим образом стимулируется. Большая часть эмбриональной
56

Рис. И-13. Связь в виде треугольника, сформированная между тремя спинальными ганглиями эмбриона in vitro. Рисунок иллюстрирует тенденцию нервной ткани формировать узоры нервных волокон (Weiss,
1967).
ткани эквипотенциальна, то есть ДНК во всех клетках тела, по существу, одна и та же. Это означает, что потенциал первоначально подавлен, находится в состоянии «репрессии». «Дерепрес-сия», или освобождение потенциальных возможностей, происходит тогда, когда появляются соответствующие условия. Определение эмбриологами того, каковы эти условия, заняло почти целое столетие. В настоящее время хорошо известно, что РНК и некоторые эндокринные секреты являются индукторами такой «дерепрессии». Маловероятно, что эти химические соединения несут в себе всю информацию, необходимую для индукции; большая доля специфики процесса должна быть отнесена за счет субстрата. Например, РНК, выделенная из печени теленка, может быть использована
57
как индуктор для хрусталика глаза крысы. Отношения между индуктором и субстратом, по-видимому, следующие.
1. Индукторы извлекают и реализуют потенциальную генетическую информацию организма
2. Индукторы в известной мере специфичны в отношении характера извлекаемой генетической информации, но они неспецифичны относительно вида особи или ткани
3. Индукторы определяют общую схему индуцированного свойства, специализация деталей возникает в результате деятельности субстрата
4. Индукторы непосредственно не являются толчком для развития, они составляют особый класс стимулов
5. Чтобы быть эффективными, индукторы должны взаимодействовать с субстратами Однако, чтобы вызвать эффект, недостаточно простого контакта — ткань должна быть готова правильно реагировать
6 Индукция обычно развивается в результате действия в двух направлениях благодаря химическому взаимодействию между индуктором и субстратом.
Данные о роли РНК в сохранении следов памяти в настоящее лремя могут быть объяснены при помощи модели, основанной на
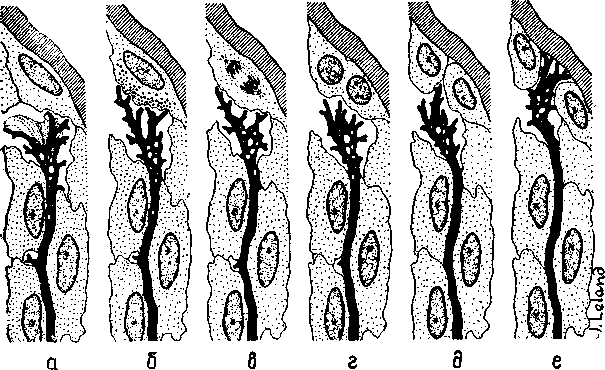
Рис. И-14 Шесть фаз гипотетического процесса индукции
этом эмбриональном процессе индукции. Модель предполагает, что возбуждение нервных структур сопровождается образованием РНК. Эта нейронная РНК вызывает изменения в окружающей «лигодендроглии, в результате чего начинается химический процесс между нейроном и глией, характеризующийся реципрокными отношениями изменений нейронной и глиальной концентрации РНК (и множества метаболитов). Происходит изменение в функциональном взаимодействии между глией и нейроном. На первом этапе может возникнуть соответствующее изменение концентрации РНК в глиальной клетке, которое затем в течение длитель-
53
ного времени будет вызывать изменение в структурах липидов, протеинов и липопротеинов во всех больших молекулах, составляющих мембраны, через которые осуществляется взаимодействие нейрона с глией. Такие макромолекулярные изменения могут влиять на легкость освобождения и разрушения нейронных медиаторов. Эти структурные изменения обратимы и могут стираться или быть вытеснены другими изменениями. Однако, если при неоднократном повторении одного и того же типа воздействия' изменения в молекулярной структуре будут продолжаться достаточно долго, они вызовут эффективные изменения мембранной проницаемости, которые в свою очередь способствуют появлению в возбужденном нейроне большого количества РНК, метаболитов и медиаторов, оказывающих влияние на окружающую его глию в том месте, где происходит деление глиальной клетки. Конус роста нейрона, освободившийся от инкапсулированной глии, свободно воздействует на недавно сформировавшиеся дочерние клетки в образует новые связи с нейронами. Таким образом, клеточное деление олигодендроглии направляет конусы роста нейронов центральной нервной системы во многом так же, как на периферии их рост направляют соответствующие шванновские клетки (рис. П-14).
КОНСОЛИДАЦИЯ СЛЕДОВ
Любое изменение глиально-нейронной организации следует рассматривать как изменение в микроструктуре активности медленных потенциалов соединений. Как мы уже говорили, такие изменения могут быть временными либо вести к возникновению более устойчивых изменений в нервной системе, связанных с ростом нейронов. Имеется много данных, свидетельствующих о том,, что для фиксации в мозгу следов памяти — энграмм — требуется время. Эти данные вполне согласуются с индукционной моделью хранения следов памяти, потому что они указывают на возможность выделения по крайней мере двух — если не больше — процессов в зависимости от времени их появления.
После достаточно сильной травмы головы человек не способен воспроизвести в памяти события, происходившие в период, непосредственно предшествовавший травме. Длительность этого периода зависит от тяжести травмы. Этот феномен, названный ретроградной амнезией, может быть использован для изучения фиксации следов, то есть для изучения процессов консолидации.
Самым распространенным методом, чтобы вызвать ретроградную амнезию в лабораторных условиях, является применение электроконвульсивного шока на крысах, хотя используются также и другие методы, такие, как удар по голове, быстрая анестезия с помощью эфира или введение определенных веществ.
59
вызывающих амнезию. Полученные данные говорят о том, что, чем скорее после того, как возник навык, вызываются конвульсии, тем сильнее их тормозное, амнестическое действие на последние из навыков, приобретенные в этом эксперименте. Так, в одном исследовании (Chorover and Schiller, 1965) максимальный эффект отмечался в тех случаях, когда конвульсии следовали не позднее, чем через 15 сек после условной реакции избегания. Эффект практически исчезал, когда проверку выработки условного рефлекса и конвульсии разделял интервал длительностью 1 час. Эффект в значительной мере зависит от сложности задания, которое нужно было выполнить.
Однако это еще не все. При соответствующих условиях возникает более сложный, двухэтапный процесс консолидации. Допустим, испытуемый должен нажимать на рычаг, чтобы получить в качестве вознаграждения пищу, предъявляемую со слегка меняющимися интервалами. В какой-то момент выполнения задачи включается сигнал, за которым через определенный промежуток времени постоянно следует удар электрического тока в ногу. Как правило, при сигнале двигательный ответ испытуемого значительно уменьшается или совсем исчезает. Было высказано предположение, что этот эффект вызывается возникновением определенного эмоционального состояния. После действия тока и выключения сигнала восстанавливается нормальная скорость ответа. Когда подобный электрошок давался через 48 часов после последнего опыта и испытание возобновилось через четыре дня после завершения серий с электрораздражением, все испытуемые, подвергавшиеся его действию, были совершенно не способны «эмоционально» реагировать на сигнал. С другой стороны, если повторный опыт отодвигался на 30, 60 или 90 дней после завершения серий с электрораздражением, снова был получен условный эмоциональный ответ. В этих экспериментах (Brady, 1951) превышался критический интервал, в течение которого может быть максимально нарушен процесс консолидации, однако их результаты можно интерпретировать и как указание на то, что в течение одного-двух дней после эксперимента оказывается ослабленным какой-то механизм, необходимый для воспроизведения следа, на основе которого возникает эмоциональная реакция.
Результаты этих экспериментов по консолидации следов памяти можно интерпретировать, в соответствии с индукционной моделью, следующим образом: сразу же после испытания появление шоковых конвульсий, анестезия и т. д. препятствуют метаболическому обмену, происходящему между нейроном и олигодендрог-лией, тормозя индукцию, вызываемую путем секреции РНК в нерве и сопровождающуюся ее активацией в глии. Следовательно, образование энграммы прекращается и припоминание становится невозможным. Напротив, когда конвульсии возникают позднее, связанная с РНК фаза процесса индукции оказывается уже завер-
60
шенной. В этом случае временно приостанавливается деление глиальной клетки, амебовидные движения окончаний нейронов временно прекращаются и конус роста втягивается назад. Однако при восстановлении этих механизмов процесс запечатления энг-раммы продолжается с того момента, когда он был прерван, и при завершении этого процесса становится возможным припоминание.
Чтобы твердо установить, что консолидация следа памяти действительно происходит, необходимо найти такой метод, который улучшал бы научение. Это, в сущности, нетрудно. Когда крысам вводили сульфат стрихнина или аналогичные возбуждающие вещества не ранее чем за 10 мин до начала или 30 сек спустя после пробы на прохождение лабиринта или на выработку дифференцировки, то навык у них вырабатывался быстрее, нем у контрольных; консолидация, таким образом, происходила скорее (McGaugh and Petrinovich, 1959).
Эти эксперименты служат красноречивым свидетельством того, что консолидация является необходимым условием возникновения следа памяти. Консолидация возникает в мозгу, но что же именно происходит во время этого процесса, который делает возможным запоминание? Ответ на этот вопрос могут дать следующие наблюдения.
В моей лаборатории на различные области коры мозга обезьян наносилась алюминиевая паста для создания локальных раздражений, которые в отдельных случаях вызывали эпилепсию. Эти раздражения проявлялись в измененной электрической активности — в возникновении патологических медленных волн и спайковых разрядов. Хотя такие нарушения не мешали обезьянам помнить решение задач, с которыми они неоднократно имели дело до раздражения, эти нарушения замедляли первоначальный процесс выработки данного навыка почти в пять раз (Pribram, 1966). Более того, способность вырабатывать такие навыки в целом не подвергалась нарушению; оно касалось только тех задач, выполнение которых было связано с определенной частью мозга, и ограничивалось лишь начальным периодом процесса выработки навыков, предшествующим появлению первых признаков научения. Таким образом, вызываемые в этом опыте раздражения не всегда блокируют консолидацию следов.
Может быть, одиночная энграмма, ограниченная лишь одним нейронным фокусом, является недостаточно эффективной, чтобы проявить себя? Большинство результатов экспериментов по научению у человека показывает, что, для того чтобы запомнить задание, необходимо его многократное повторение (Trabasso and Bower, 1968).
Во время повторения происходит такое распределение повторяемого материала, что он включается в целую систему связей предварительно зафиксированного опыта. Таким образом, резуль-
61
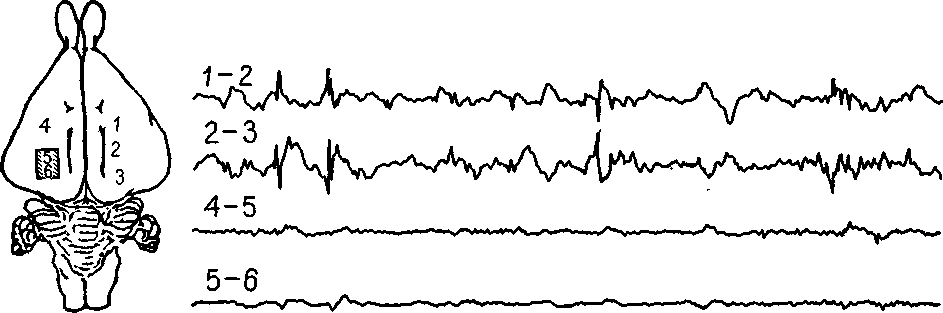
* Рис. 11-15. Спонтанные разряды во вторичном эпилептогенном очаге активности. Вторичный очаг активности (два верхних канала) продолжает разряжаться, несмотря на прекращение разрядов в области первичного разрушения (нижние два канала). Калибровка: 100 мкв и 1 сек (Morrell, 1967).
таты экспериментов с нарушениями, вызванными раздражением, можно объяснить тем, что процесс повторного возникновения и распределения энграммы был задержан. Проверку этой интерпретации можно осуществить путем сравнения результатов научения обезьян с повреждениями мозга в условиях распределенного концентрированного научения.
Гистологический анализ мозговой ткани, подвергшейся воздействию алюминиевой пасты, показывает наличие спутанных клубочков нервных волокон, во многом напоминающих соответствующие образования в периферических нервах, когда их рост не направляется соответствующей колонкой шванновских клеток. Возможно ли, чтобы это воздействие избирательно уничтожало олигодендроглию, вызывая ненаправленный рост нервных волокон? Химический анализ ткани, в которую была введена алюминиевая паста, конечно, не осуществим. Однако, чтобы получить ответ на этот вопрос, был проведен остроумный эксперимент (Morrell, 1961, рис. П-15). Разрушение, вызванное раздражением в одном полушарии головного мозга, через несколько месяцев вызывает в контралатеральной коре появление «зеркального очага» измененной электрической активности, который возникает при участии мозолистого тела. Этот «зеркальный очаг» непосредственно не испытывал разрушающего химического воздействия, но обладал всеми признаками, характерными для первичного очага разрушения, вызванного раздражением. Существенно ли меняется РНК в «зеркальном очаге» по сравнению с нормальной тканью мозга? Может ли измененная РНК быть непосредственной причиной этого раздражения и последующих трудностей в восстановлении следа памяти?
Индукция в памяти, так же как и индукция в эмбриогенезе, является, видимо, многоэтапным процессом, который требует времени для своего завершения. Можно ожидать, что каждая стадия в таком процессе имеет свое уязвимое место, которое можно выявить с помощью соответствующего метода, примененного в критический момент. Уже многое известно о «критических периодах» в развитии эмбрионов и поведения в первые годы жизни. Определение природы критических, то есть сензитивных, периодов в процессе образования следов памяти представляет собой богатую область для исследования и экспериментов.
Можно еще многое сказать — и это будет сделано — о консолидации и модели индукции, но сначала нам нужно составить более полное представление о принципах работы мозга и о структуре психологических процессов. Поэтому рассмотрим в следующей главе некоторые из способов, посредством которых жизненный опыт организма может вносить свои изменения в мозг.
63
РЕЗЮМЕ
Длительно сохраняющиеся изменения происходят в соединительных аппаратах мозговой ткани. Хотя зрелые нервные клетки не делятся, механизм постоянной модификации мозговой ткани обладает многими свойствами механизма дифференциации эмбриональной ткани. В обоих случаях можно наблюдать экспериментально вызванный направленный рост новых нервных волокон, который меняет пространственную структуру связей между нейронами. Следовательно, долговременная память является скорее функцией соединительных структур, чем функцией процессов в самой нервной клетке, генерирующей нервные импульсы.
Глава III
ЗАТУХАНИЕ АКТИВНОСТИ В НЕРВНОЙ СИСТЕМЕ И ТОРМОЗНЫЕ ВЗАИМОДЕЙСТВИЯ
ОРИЕНТИРОВОЧНАЯ РЕАКЦИЯ И ПРИВЫКАНИЕ
В первой главе были проанализированы два основных класса переменных, характеризующих деятельность мозга. Взятый в качестве модели процесс воспроизведения речи показал возможности, открывающиеся благодаря модификации этих двух переменных. Во второй главе были детально рассмотрены данные, свидетельствующие о пластичности мозга и его способности к постоянным изменениям; однако мы еще не говорили о свойствах самого механизма, который делает возможным осуществление таких изменений. При игре на скрипке, флейте или органе участвуют те же самые основные переменные, что и в речевом общении, хотя сами инструменты различаются тем, как они воспроизводят звуки.
Поэтому настоящая глава начинается с рассмотрения организации нервной системы, позволяющей ей быть аппаратом, создающим все богатство нашего опыта и поведения. Эта организация во многом зависит от относительно постоянных структур, образующихся в тех системах, посредством которых мозг приходит в соприкосновение со своим окружением. Благодаря своей доступности эти системы могут служить некоторыми частными моделями и, следовательно, подвести к исследованию более центральных процессов, анализ которых требует иногда больших усилий. Поскольку довольно сложная организация имеет место уже при первоначальном контакте организма с его окружением, уместно начать наше исследование именно с этого процесса.
Экспериментальные данные говорят о том, что в любой момент времени происходит отбор сенсорного возбуждения с помощью хранящейся в памяти информации о прошлом опыте. Этот процесс сравнения, в ходе которого выявляется, соответствует ли текущее возбуждение хранящимся в мозгу записям, направляет как внимание человека, так и его действия.
Е. Н. Соколов (1960) провел следующий простой опыт, в котором обнаружилось одно из тех противоречий, которые порождают