
 Авторефераты по всем темам >>
Авторефераты по биологии
Авторефераты по всем темам >>
Авторефераты по биологии
Рекомбинантные цитокины и цитокин-связывающие белки
Автореферат докторской диссертации по биологии
|
Страницы: | 1 | 2 | 3 | 4 | 5 | 6 | |
 СКАЧАТЬ ОРИГИНАЛ ДОКУМЕНТА
СКАЧАТЬ ОРИГИНАЛ ДОКУМЕНТАРабота выполнялась в ФБУН ГН - ВБ Вектор в рамках научных тем организации, по гранту Международного научно-технического центра #1685 Разработка нового терапевтического препарата на основе ортопоксвирусного белка, связывающего фактор некроза опухолей человека, по грантам РФФИ 06-04-48074а и 10-04-00387а, в рамках Федеральной целевой программы Научные и научно-педагогические кадры инновационной России, госконтракт № 100430/--3052/266.
Конструирование рекомбинантных плазмид для экспрессии природных и синтетических генов в клетках E. coli выполнено автором лично и в соавторстве с В.В. Кравченко, Т.С. Бондарь (НИКТИ БАВ, г. Бердск) и В.Н. Добрыниным, С.А. Филипповым, В.Г. Коробко (ГУН Институт биоорганической химии им. М.М. Шемякина и Ю.А. Овчинникова, г. Москва). Физико-химический анализ рекомбинантных белков hIFNa-2b и iGM-CSF выполнен автором лично и в соавторстве с З.А. Акименко, СИ. Ястребовым, В.И. Офицеровым (ЗАО Вектор-Бест, г. Новосибирск). Изучение свойств штаммов-продуцентов E.coli SG20050/pIF16 и E.coli SG20050/p280GM проведено в соавторстве с И.А. Андреевой (ФБУН ГН - ВБ Вектор,) и Н.В. Наумовой (ЗАО Вектор-Медика, р.п. Кольцово). Выделение рекомбинантных белков hIFNa-2b, iGM-CSF и MuTNF проведено в соавторстве с Л.Р. Лебедевым, В.П. Романовым, Н.М. Пустошиловой (ФБУН ГН - ВБ ВЕКТОР, р.п. Кольцово), А.Н. Закабуниным и Ю.Н. Козловой (ГУН ИХБФМ СО РАН, г. Новосибирск). Изучение биологических свойств рекомбинантного iGM-CSF проведено в соавторстве с СК. Халдояниди, И.А. Орловской, Л.Б. Топорковой, Л.В. Сахно, СВ. Сенниковым (ГУН НИИКИ СО РАМН, г. Новосибирск). Изучение СгтВ-белков ортопоксвирусов выполнено автором под общим руководством СН.а ааЩелкунова.
9
Конструирование рекомбинантных бакуловирусов BTRI67, BTRz96 и
BTRgr90 проведено И. А. Рязанкиным, З.А. Максютовым, A.B.
Тотмениным при участии автора. Компьютерный анализ
аминокислотных последовательностей СгтВ-белков проведен A.B.
Тотмениным и Д.В. Антонецом (ФБУН ГН - ВБ ВЕКТОР, р.п.
Кольцово). Выделение СгтВ-белков проведено Л.Р. Лебедевым (ФБУН
ГН - ВБ ВЕКТОР, р.п. Кольцово). Изучение физико-химических и
иммунологических свойств СгтВ-белков проведено автором лично и в
соавторстве с И.А. Рязанкиным, Н.М. Пустошиловой и Г.Н.
Афиногеновой (ФБУН ГН - ВБ ВЕКТОР, р.п. Кольцово). Изучение
TNF-нейтрализующих эффектов СгтВ-белков на клетках линии L929 и
на модели ЛПС-индуцированного септического шока проведено автором
лично, Т.С. Непомнящих под руководством автора и совместно с Г.В.
Кочневой и A.A. Гражданцевой (ФБУН ГН - ВБ ВЕКТОР, р.п.
Кольцово). Изучение TNF-нейтрализующих эффектов белка VARV-
CrmBаа ваа экспериментальныха моделяха костномозгового
колониеобразования и ингибирования миграции клеток Лангерганса выполнено совместно с Л.Б. Топорковой, И.А. Орловской, Е.А. Вязовой (ГУН НИИКИ СО РАМН, г. Новосибирск). Автор также приносит благодарность настоящим и бывшим сотрудникам ФБУН ГН - ВБ ВЕКТОР, внесшим свой вклад в проведение настоящего исследования - В.А. Лихошваю, В.В. Шамину, О.И. Серпинскому, Е.И. Рябчиковой, Е.М. Малковой, И.В. Виноградову, В.А. Агеенко и сотрудникам отдела Геномных исследований и разработки методов ДНК-диагностики поксвирусов. Особую благодарность автор выражает к.х.н. В.В. Кравченко - инициатору изучения механизмов экспрессии генов в составе полицистронных оперонов и Е.А. Каргиновой за радость совместной работы, д.б.н., профессору С.К. Василенко, отношение которого к жизни и науке являются примером.
10
По теме диссертации опубликовано 20 печатных работ (16 статей в научных журналах и сборниках научных трудов). Часть результатов защищена патентами (4 Патента РФ). Патент РФ № 2241754 включен в список 100 лучших патентов России за 1997-2007 гг.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ И ИХ ОБСУЖДЕНИЕ
1. Конструирование рекомбинантных плазмид для экспрессии в клетках E.coli природных и синтетических генов в составе искусственных полщистронных оперонов
Взаимосвязанная трансляция генов бактериальных оперонов
позволяет предположить целесообразность использования этого принципа при конструировании экспрессионных плазмид. В работах (Кравченко В.В. и др., 1987, 1988) показано, что наиболее эффективная экспрессия полусинтетического гена lacZ, находящегося в составе искусственного полицистронного оперона, наблюдается, когда его последовательность SD перекрывается или находится в непосредственной близости с терминирующим кодоном предшествующего гена B-белка фага М13. Высокий уровень экспрессии дистального гена оперона достигается за счет эффективной инициации трансляции, которая становится возможной благодаря разрушению вторичной структурыа мРНК, происходящему ва процессе трансляции
1 +аа 9+
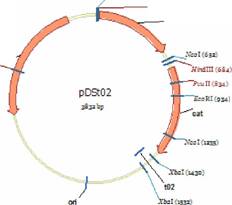
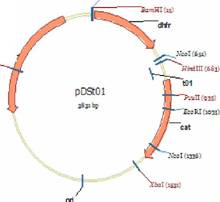
проксимальногоаа гена.аа Рекомбинантныеаа плазмидыаа pDStoа иаа pDSto (Stueber D., Bujard H., 1982), физические карты которых представлены на рис. 1, использовали в качестве исходных векторных молекул.
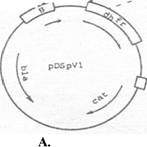
Клонированием PstI/EcoRI-фрагмента плазмиды pSK-381 (Серпинский и др., 1982), содержащего начало гена bla, промотор ???? и ген белка оболочки (B-белка) фага М13тр7 без 4-х С-концевых ко донов, и синтетического EcoRI/BamHI-адаптера в плазмиде pDSto получена плазмида pDSpVl, физическая карта которой представлена на рис. 2А. Эта плазмида представляет искусственный полицистронный оперон, в котором область стыка генов B-белка и dhfr организована таким образом,
11
что SD-последовательность дистального гена оперона перекрывается с терминирующим кодоном проксимального, как это изображено на рис.
ЗА.
|
|
'dliFr |
1>лаГггамаа ---: hнн |
b-лаггамаа |
Рис. 1 Физическая карта плазмид pDSto1+ и pDSto2+. bla-, dhfr- cat- гены Я-лактамазы, дигидрофолатредуктазы и хлорамфениколацетилтрансферазы, соответственно; ori - область начала репликации; tO - терминатор транскрипции фага ?. Указано положение сайтов узнавания эндонуклеаз рестрикции Xhol, EcoRl BamHT HindlII, Ncol, Xbal и Pstl.
ifn oZ
,AЫН!AAЬЬAT..ATGTC. |
dhfr .TTCACO.. .TAAACTUАЛПТАпОТПССОСАТСАТааТ
SeoRI |
жжжХ^аа "Г~аа ~~Г~аа ?_ ж* "
Banni . |
^ -.а SD
 EcoRОаа Ч -
EcoRОаа Ч -
Li+
Рис. 2 Физические карты плазмид pDSpVl (А) и pIF6/8 (Б). Указаны гены bla, dhfr, cat и ifria-2; терминатор транскрипции to фага ?, полилинкер (pl), а также положение сайтов узнавания эндонуклеаз рестрикции EcoRI и BamHI. Подчеркнуты снизу терминирующие кодоны, сверху - перекрывание последовательности SD и кодона терминации.
В качестве дистального гена в плазмиде pDSpVl выступает природный ген dhfr. Для определения эффективности экспрессии синтетического гена интерферона альфа-2Ь человека в составе искусственного полицистронного оперона сконструированы рекомбинантные плазмиды pIF6/8, p381IFN и p280IFN. Источник гена ifna-2b - рекомбинантная плазмида pLelFlac (Колосов М.Н. и др.,
12
1984). Нуклеотидная последовательность искусственного гена ifnг-2 и кодируемая им аминокислотная последовательность представлены на рис. 4.
А.
----- В-белок---------------------- ->
SDdhfr-^---- dhfr
... GAATTCACCTAACTGAAGTAAGGATCCGCATCATGGTT.........
2а 1а О
Б.
SD,fДa2<rЧifrw2-Ч->аа SDdhfr<....dhfr
TAACTGAAGTAAGGATCAATTCTA JGTGT...TAG. 3 5n... GGATCCGC ATC4 JGGTT...
2а 1а 0 Рис. 3 Последовательность нуклеотидов в области стыка генов В белка и dhfr (А); В белка и ifria-2а (Б). Курсивом обозначены терминирующие кодоны гена белка В и инициирующие кодоны генов dhfr и ifria-2.а Жирным шрифтом обозначены SD-последовательности генов; n - нуклеотидный остаток.
А. (Колосов М.Н. и др., 1984)
ATGTGTGATCTGCCGCAGACTCACTCTCTGGGTTCTCGTCGTACTCTGATGCTGCTGGCTCAGATGCGTCGTA
TCTCTCTTTTCTCЮTGCCTGAAAGACCGЮCATGACTTCGGЮTTCCCGCAGGAAGAGTTCGGTAACCAGTTCCA
GAAAGCTGAAACTATCCCTGTTCTGCATGAAATGATCCAGCAAATCTTCAACCTGTTCTCTACTAAAGACTCT
TCTGCTGCTTGGGACGAAACCCTGCTGGACAAATTCTACACTGAACTGTACCAGCAACTCAACGACCTCGAGG
CTTGTGTTATTCAGGGCGTTGGTGTTACCGAAACTCCGCTGATGAAAGAGGATAGCATCCTGGCTGTTCGCAA
GTATTTCCAGCGTATCACTCTTTACCTGAAGGAAAAGAAGTATAGCCCGTGTGCTTGGGAAGTTGTTCGTGCT
GAGAT CAT GC GT T С СТ T СТ С С CЮGT CTACTAACCGCaGGAATCTCTGCGTTCTAAAGAATAG
>, frApe+1, 166 bases
MCDLPQTHSLGSRiCTLMLLAQMRRISLFSCLKDRHDFGFPQEEFGNQFQK AETIPVLHEMIQQIFNLFSTKDSSAAWDETLLDKFYTELYQQLNDLEACV IQGVGVTETPLMKEDSILAVRKYFQRITLYLKEKKYSPCAWEWRAEIMR SFSLSTNLQESLRSKE*
Б. GenBank: AY255838.1
tgtgacctaccacaaacccacagcctgggtagcaggaggaccttgatgctcctggcgcagatgaggagaatct
ctcttttctcctgcttgaaggacagacatgactttggatttccccaggaggagtttggcaaccagttccaaaa
ggctgaaaccatccctgtcctccatgagatgatccagcagatcttcaacctcttcagcacaaaagactcatct
gctgcttgggatgagaccctcctagacaaattctacactgaactctaccagcagctgaatgacctggaagcct
gtgtgatacagggggtgggggtgacagagactcccctgatgaaggaggactccattctggctgtgaggaaata
cttccaaagaatcactctctatctgaaagagaagaaatacagcccttgtgcctgggaggttgtcagagcagaa
atcatgagatctttttctttgtcacaaacttgcaagaaagtttaagatctaaagagtaatga
>frApe+l, 165 bases
CDLPQTHSLGSRiCTLMLLAQMRRISLFSCLKDRHDFGFPQEEFGNQFQK AETIPVLHEMIQQIFNLFSTKDSSAAWDETLLDKFYTELYQQLNDLEACV IQGVGVTETPLMKEDSILAVRKYFQRITLYLKEKKYSPCAWEWRAEIMR SFSLSTNLQESLRSKE*
Рис. 4 Нуклеотидные последовательности искусственного (А) и природного (В) генов ifriOL-2b и кодируемые ими аминокислотные последовательности IFNa-2b. Подчеркнуты нуклеотиды искусственного гена, отличающиеся от природного. Сайт узнавания эндонуклеазы рестрикции Pstl выделен жирным курсивом. Жирным шрифтом выделены аминокислотные остатки R23, Н34 позволяющие типировать синтетический ген как ifria-lh.
13
Физическая карта плазмиды pIF6/8 и последовательность нуклеотидов в области стыка генов B-белка и ifna-2b представлены на рис. 2Б и рис. ЗБ, соответственно. В этой плазмиде ген dhfr уже не входит в полицистройный оперон.
При конструировании плазмиды p381IFN с помощью синтетического олигонуклеотидного адаптера, структура которого представлена на рис. 5, изменили структуру гена ifna-2b, что не привело к изменению соответствующей аминокислотной последовательности. В результате реконструкции одна из замен G-^A приводит к утрате сайта Pstl в гене интерферона, другая изменяет терминирующий кодон TAG-^TAA. Также при встройке адаптера происходит утрата сайта узнавания EcoRI-934 плазмиды pDSto , что позволяет заменой PstI/EcoRI-фрагмента плазмиды p381IFN на соответствующий фрагмент, содержащий тандем промоторов и часть лидерного пептида триптофанового оперона.со//, получить плазмиду p280IFN.
EcoRITSD -пifnPstl*EcoRI*
CGAATTCGACA....... ACTGAGGATC... ATGTCTGA... .CCrGCЖ4TCTCTCCGTTCTAAAGAATAAжATTЖ47TC
Gаа G
~pDS381 -><--------------------------- pIF6/S-------------------- -> <----------------------- -A------------------------------------ ->
Рис.а 5 Отличия структуры гена ifna-lh в плазмиде p381IFN от его структуры в плазмиде pIF6/8.
2.аа Эффективность экспрессии генов в составеа искусственных полицистронных оперонов Полученными плазмидами pDSpVl, pIF6/8, p381IFN и p280IFN
трансформировали клетки E.coli SG20050 и индивидуальные AprTcr-
трансформанты использовали для получения клеточных лизатов,
которые анализировали с помощью электрофореза в 12% SDS-ПААГ и
методом радиоиммуноанализа (РИА). Результаты электрофоретического
фракционирования суммарных лизатов клеток E.coli SG20050/pDSpY\ и
SG20050/nl?6/8 представлены на рис. 6. Плазмиды pDSpVl и pIF6/8, по
сравнению с контрольной плазмидой pLelFlac, детерминируют синтез
новыхаа полипептидов, составляющих мажорныеаа полосы н фоне
14
остальных клеточных белков, которые по молекулярным массам соответствуют дигидрофолатредуктазе мыши (22 кДа) и альфа-2 интерферону человека (кДа), соответственно.
12аа 3аа 4
1-1
Ol
Рис. 6 Фракционирование лизатов клеток E.coli SG20050/pDSpVl (2) и E.coli SG20050/pWm (4) в 12% SDS-ПААГ. 1 - лизат клеток E.coli SG20050/pLeIFlac; 3-химотрипсиноген (25 кДа) и миоглобин (17 кДа), соответственно.
Плазмида pIF6/8 позволяет количественно оценить уровень экспрессии генов искусственного бицистронного оперона путем измерения количеств белков В и IFNa-2b, синтезирующихся в клетках E.coli, несущих эти плазмиды. Результаты РИА представлены в табл. 1.
Таблица 1
Определение количеств белка В и лейкоцитарного интерферона а-2Ь человека в лизатах клеток E.coli SG20050/pDSpVl и E.coli С20050/р1?Ш.
Плазмида |
Белок В (пмоль/109 клеток) |
Интеферон ос-2Ь (пмоль/109 клеток) |
pDSpVl |
28 |
0 |
pLelFNlac |
0 |
1.5 |
pIF6/8 |
32 |
1600 |
Можно видеть, что уровень экспрессии дистального гена (ifna-2b) практически в 50 раз превышает уровень экспрессии проксимального (VIII), т.е. наблюдается эффект усиления экспрессии целевого гена в результате трансляционного сопряжения.
Экспрессионныеа свойства плазмида pIF6/8,а p381IFN иа p280IFN исследовали определением процентного содержания IFNoc-2b в клетках
15
E.coliа денситометрией.а Результатыа проведенныха экспериментов представлены в табл. 2.
3. Экспрессия дуплицированного гена ifna-2b в составе полицистронных оперонов с различным транскрипционным контролем
Увеличение в клетке количества копий определенного гена вызывает повышение уровня его продукта. Представляет интерес исследование экспрессии повторяющихся генов, находящихся в составе полицистронного оперона. Для направленной дупликации генов ifna-2b Pstl-фрагмент плазмиды pLelFlac, содержащий начало гена bla, 5'-концевую часть ifna-2b, PstI/EcoRI-адаптор в виде комплементарных дезоксиолигонуклеотидов длиной 30 и 38 п.о. и PstI/EcoRI-фрагмент плазмиды pDSto использовали для получения промежуточной плазмиды p95IFN. Для получения тандема генов ifna-lh Pstl/EcoRI- фрагменты плазмид p95IFN и p381IFN в присутствии адаптера лигировали с интерферон-содержащими Pstl-фрагментами плазмид pLelFNlac и pIF6/8, получая в результате плазмиды p95/2IFN и p381/2IFN, соответственно. Заменой Pstl/EcoRI фрагмента плазмиды p381/2IFN на соответствующий Pstl/EcoRI фрагмент плазмиды pDS280 получили плазмиду p280/2IFN. На рис. 7 схематически изображена структурно-функциональная организация полицистронных оперонов под контролем промоторов Piac (p95/2IFN; ifna-ifna), ???? (p381/2IFN; IX-VIII-ifna-ifna) и P^ (p280/2IFN; L-ifna-ifna).
Клетки E.coli SG20050 трансформировали плазмидами pIF6/8, p381IFN, p280IFN, p95/2IFN, p381/2IFN и p280/2IFN и определяли в суммарных лизатах клеток уровень синтеза IFNoc-2b денситометрией. Относительный уровень синтеза интерферона представлен в табл. 3, а результаты фракционирования суммарных лизатов плазмидосодержащих клеток - на рис. 8.
16
p95/2IFN
-AGGБ-ЧUnЧATG__________ ifnaTAAGGKЧ8nЧ-ATG_________ iЯ a______ TAA
p381/2IFN
ЧIX--------- ATGA------- VIII------ rЖ4GGAЧ8nЧATG_________ ifhaTAAGGAЧ8nЧ-
A TG_ifn a___ TAAЧ
p280/2IFN
ЧtrpL ЧrЖ4GGAЧ8nЧATG__________ ifnaTAAGGKЧ8nЧ-ATG_________ iЯ a______ TAAЧ
pIF16
ЧtrpL ЧrЖ4GGAЧ8nЧATG__________ ifnaTAAGGKЧ69nЧ-A TG________ iЯa______ TAAЧ
Рис. 7 Структурно-функциональная организация сигналов инициации трансляции генов интерферона в составе полицистронных оперонов плазмид p95/2IFN, p381/2IFN, p280/2IFN и pIF16 (раздел 5). Инициирующие и терминирующие кодоны генов iЯi(x-2b выделены жирным курсивом, последовательности TSD (TAAGGA) -подчеркнуты. Цифрами обозначено расстояние в нуклеотидных остатках между соответствующими регуляторными сигналами.
|
Страницы: | 1 | 2 | 3 | 4 | 5 | 6 | |
Авторефераты по всем темам >>
Авторефераты по биологии